ผู้เชี่ยวชาญทางการแพทย์ของบทความ

สิ่งตีพิมพ์ใหม่
ไฮโปทาลามัส
ตรวจสอบล่าสุด: 07.07.2025

เนื้อหา iLive ทั้งหมดได้รับการตรวจสอบทางการแพทย์หรือตรวจสอบข้อเท็จจริงเพื่อให้แน่ใจว่ามีความถูกต้องตามจริงมากที่สุดเท่าที่จะเป็นไปได้
เรามีแนวทางการจัดหาที่เข้มงวดและมีการเชื่อมโยงไปยังเว็บไซต์สื่อที่มีชื่อเสียงสถาบันการวิจัยทางวิชาการและเมื่อใดก็ตามที่เป็นไปได้ โปรดทราบว่าตัวเลขในวงเล็บ ([1], [2], ฯลฯ ) เป็นลิงก์ที่คลิกได้เพื่อการศึกษาเหล่านี้
หากคุณรู้สึกว่าเนื้อหาใด ๆ ของเราไม่ถูกต้องล้าสมัยหรือมีข้อสงสัยอื่น ๆ โปรดเลือกแล้วกด Ctrl + Enter
ไฮโปทาลามัสเป็นส่วนล่างของไดเอนเซฟาลอนและมีส่วนร่วมในการสร้างพื้นของโพรงสมองที่ 3 ไฮโปทาลามัสประกอบด้วยไคแอสมาของเส้นประสาทตา เส้นประสาทตา ปุ่มสีเทาที่มีกรวย และส่วนเต้านม
ไคแอสมาออปติก (chiasma opticum) เป็นสันขวางที่เกิดจากเส้นใยของเส้นประสาทตา (เส้นประสาทสมองคู่ที่ 2) ข้ามไปบางส่วนทางด้านตรงข้าม (เกิดการไขว้กัน) สันนี้ต่อเนื่องไปด้านข้างและด้านหลังในแต่ละด้านเข้าสู่ช่องประสาทตา (tratus opticus) ช่องประสาทตาตั้งอยู่ตรงกลางและด้านหลังจากสารที่มีรูพรุนด้านหน้า โค้งไปรอบก้านสมองจากด้านข้าง และสิ้นสุดด้วยรากสองรากในศูนย์การมองเห็นใต้เปลือกสมอง รากข้างที่มีขนาดใหญ่กว่า (radix lateralis) จะเข้าใกล้กับ lateral geniculate body และรากกลางที่บางกว่า (radix medialis) จะไปที่ superior colliculus ของหลังคาของสมองกลาง
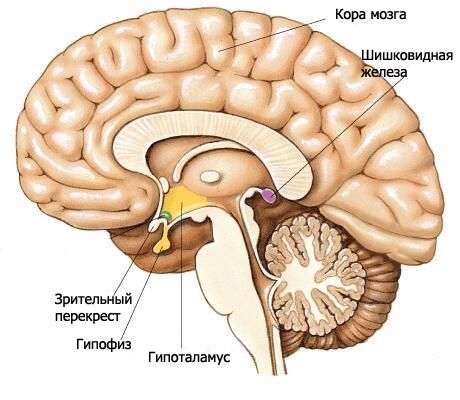
แผ่นปลายประสาทซึ่งเป็นส่วนหนึ่งของเทเลนเซฟาลอนนั้นอยู่ติดกับพื้นผิวด้านหน้าของไคแอสมาของเส้นประสาทตาและรวมเข้ากับไคแอสมา แผ่นปลายประสาทนี้ปิดส่วนหน้าของรอยแยกตามยาวของสมองและประกอบด้วยชั้นบางๆ ของเนื้อเทา ซึ่งในส่วนด้านข้างของแผ่นประสาทนั้นจะต่อเนื่องเข้าไปในสารของกลีบหน้าผากของซีกสมอง
ด้านหลังไคแอสมาของเส้นประสาทตาคือตุ่มสีเทา (tuber cinereum) ซึ่งอยู่ด้านหลังมีต่อมน้ำนม และด้านข้างเป็นเส้นประสาทตา ด้านล่างตุ่มสีเทาจะผ่านเข้าไปในกรวย (infundibulum) ซึ่งเชื่อมต่อกับต่อมใต้สมอง ผนังของตุ่มสีเทาเกิดจากแผ่นเนื้อสีเทาบางๆ ที่มีนิวเคลียสของตุ่มสีเทา (nuclei tuberales) จากด้านข้างของโพรงของโพรงสมองที่ 3 รอยบุ๋มแคบของกรวยจะยื่นเข้าไปในบริเวณของตุ่มสีเทาและเข้าไปในกรวยมากขึ้น
เนื้อเยื่อเต้านม (corpora mamillaria) อยู่ระหว่างปุ่มสีเทาด้านหน้าและเนื้อเยื่อที่มีรูพรุนด้านหลัง เนื้อเยื่อเหล่านี้มีลักษณะเป็นก้อนกลมสีขาวขนาดเล็ก 2 ก้อน แต่ละก้อนมีเส้นผ่านศูนย์กลางประมาณ 0.5 ซม. เนื้อเยื่อสีขาวจะอยู่ภายนอกเนื้อเยื่อเต้านมเท่านั้น ภายในเนื้อเยื่อสีเทาซึ่งแยกนิวเคลียสส่วนกลางและส่วนด้านข้างของเนื้อเยื่อเต้านม (nuclei corporis mamillaris mediales et laterales) ออกจากกัน คอลัมน์ของ fornix จะอยู่ปลายสุดที่เนื้อเยื่อเต้านม
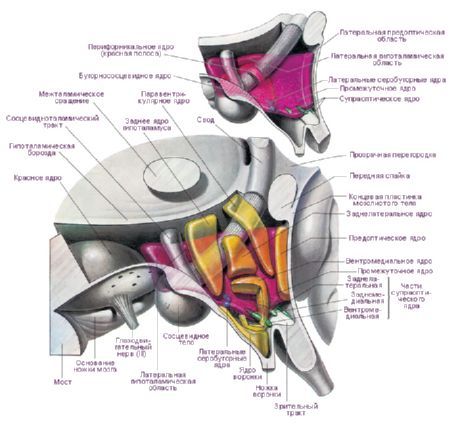
ในไฮโปทาลามัสมีบริเวณไฮโปทาลามัสหลัก 3 แห่ง ซึ่งเป็นกลุ่มเซลล์ประสาทที่มีรูปร่างและขนาดต่างกัน ได้แก่ เซลล์ด้านหน้า (regio hypothalamica anterior) เซลล์ตรงกลาง (regio hypothalamica intermedia) และเซลล์ด้านหลัง (regio hypothalamica posterior) กลุ่มเซลล์ประสาทในบริเวณเหล่านี้ประกอบกันเป็นนิวเคลียสมากกว่า 30 นิวเคลียสของไฮโปทาลามัส
เซลล์ประสาทของนิวเคลียสไฮโปทาลามัสมีความสามารถในการผลิตสารคัดหลั่ง (neurosecretion) ซึ่งสามารถส่งไปยังต่อมใต้สมองได้โดยผ่านกระบวนการของเซลล์เหล่านี้ นิวเคลียสดังกล่าวเรียกว่า นิวเคลียส neurosecretory ของไฮโปทาลามัส ในบริเวณด้านหน้าของไฮโปทาลามัสมีนิวเคลียส supraoptic (supraoptic) (nucleus supraopticus) และนิวเคลียส paraventricular (nuclei paraventriculares) กระบวนการของเซลล์ในนิวเคลียสเหล่านี้ก่อตัวเป็นกลุ่มไฮโปทาลามัส-ต่อมใต้สมอง ซึ่งสิ้นสุดที่กลีบหลังของต่อมใต้สมอง ในกลุ่มนิวเคลียสของบริเวณด้านหลังของไฮโปทาลามัส นิวเคลียส medial และ lateral ของ mammillary body (nuclei corporis mamillaris mediales et laterales) และนิวเคลียสไฮโปทาลามัสด้านหลัง (nucleus hypothalamicus posterior) เป็นกลุ่มที่ใหญ่ที่สุด กลุ่มของนิวเคลียสของบริเวณไฮโปทาลามัสระดับกลาง ได้แก่ นิวเคลียสไฮโปทาลามัสอินเฟอโรมีเดียลและซุปเปอร์เมเดียล (nuclei hypothalamic ventromediales et dorsomediales), นิวเคลียสไฮโปทาลามัสด้านหลัง (nucleus hypothalamicus dorsalis), นิวเคลียสของอินฟันดิบูลาริส (nucleus infundibularis), นิวเคลียสหัวสีเทา (nuclei tuberous) ฯลฯ
นิวเคลียสของไฮโปทาลามัสเชื่อมต่อกันด้วยระบบทางเดินรับและส่งกลับที่ค่อนข้างซับซ้อน ดังนั้น ไฮโปทาลามัสจึงมีผลในการควบคุมหน้าที่ต่างๆ ของร่างกาย การหลั่งของนิวเคลียสของไฮโปทาลามัสสามารถส่งผลต่อการทำงานของเซลล์ต่อมใต้สมอง ทำให้การหลั่งฮอร์โมนหลายชนิดเพิ่มขึ้นหรือลดลง ซึ่งจะไปควบคุมการทำงานของต่อมไร้ท่ออื่นๆ ต่อไป
การมีอยู่ของการเชื่อมต่อของระบบประสาทและของเหลวระหว่างนิวเคลียสไฮโปทาลามัสและต่อมใต้สมองทำให้สามารถรวมทั้งสองเข้าเป็นระบบไฮโปทาลามัส-ต่อมใต้สมองได้
การศึกษาด้านวิวัฒนาการทางพันธุกรรมได้แสดงให้เห็นว่าไฮโปทาลามัสมีอยู่ในคอร์ดาตาทั้งหมด ได้รับการพัฒนาอย่างดีในสัตว์สะเทินน้ำสะเทินบก และยิ่งกว่านั้นในสัตว์เลื้อยคลานและปลา นกได้แสดงความแตกต่างของนิวเคลียสอย่างชัดเจน ในสัตว์เลี้ยงลูกด้วยนม เนื้อเทาได้รับการพัฒนาอย่างมาก โดยเซลล์ของเนื้อเทาจะแยกความแตกต่างออกเป็นนิวเคลียสและฟิลด์ ไฮโปทาลามัสของมนุษย์ไม่แตกต่างจากไฮโปทาลามัสของสัตว์เลี้ยงลูกด้วยนมชั้นสูงมากนัก
มีการจำแนกนิวเคลียสไฮโปทาลามัสจำนวนมาก E. Gruntel ระบุคู่ของนิวเคลียส 15 คู่ ได้แก่ W. Le Gros Clark - 16 คู่, H. Kuhlenbek - 29 คู่ การจำแนกที่ใช้กันอย่างแพร่หลายที่สุดคือของ W. Le Gros Clark ใน Bogolepova (1968) เสนอให้แบ่งนิวเคลียสไฮโปทาลามัสออกเป็นสี่ส่วนโดยอาศัยการจำแนกข้างต้นและคำนึงถึงข้อมูลออนโทเจเนซิส:
- ส่วนหน้า หรือ ส่วนหน้า (รวมพื้นที่พรีออปติกและกลุ่มด้านหน้า - W. Le Gros Clark) - พื้นที่พรีออปติกด้านในและด้านข้าง นิวเคลียสซูพราไคแอสมาติก นิวเคลียสซูพราออปติก นิวเคลียสพาราเวนทริคิวลาร์ พื้นที่ไฮโปทาลามัสด้านหน้า
- ส่วนตรงกลางของสมอง - นิวเคลียสเวนโตรมีเดียล, นิวเคลียสดอร์โซมีเดียล, นิวเคลียสอินฟันดิบูลาร์, บริเวณไฮโปทาลามัสส่วนหลัง;
- ส่วนข้างตรงกลาง - พื้นที่ไฮโปทาลามัสด้านข้าง, นิวเคลียสไฮโปทาลามัสด้านข้าง, นิวเคลียสทูเบอโรลาเทอรัล, นิวเคลียสทูเบอโรแมมมิลลารี, นิวเคลียสรอบฟอร์นิคัล
- ส่วนหลัง หรือ ส่วนทรวงอก - นิวเคลียสทรวงอกด้านใน นิวเคลียสทรวงอกด้านข้าง
การเชื่อมต่อทางกายวิภาคของไฮโปทาลามัสยังชี้แจงความสำคัญ (ในเชิงหน้าที่) ของไฮโปทาลามัสอีกด้วย ในบรรดาเส้นทางรับความรู้สึกที่สำคัญที่สุด สามารถแยกแยะได้ดังต่อไปนี้:
- มัดสมองส่วนหน้าส่วนกลาง ซึ่งส่วนข้างเชื่อมระหว่างไฮโปทาลามัสกับหลอดรับกลิ่นและปุ่มรับกลิ่น บริเวณเพอริอะมิกดาลอยด์และฮิปโปแคมปัส และส่วนกลางเชื่อมระหว่างเซปตัม บริเวณทแยง และนิวเคลียสคอเดต
- แถบปลายซึ่งวิ่งจากต่อมทอนซิลไปยังส่วนหน้าของไฮโปทาลามัส
- เส้นใยที่ผ่านฟอร์นิกซ์จากฮิปโปแคมปัสไปยังลำตัวเต้านม
- การเชื่อมต่อระหว่างทาลามัส สไตรโอ และพาลลิโดไฮโปทาลามัส
- จากก้านสมอง - เส้นประสาทเทกเมนทัลส่วนกลาง
- จากเปลือกสมอง (เบ้าตา, ขมับ, ข้างขม่อม)
ดังนั้น แหล่งหลักของการรับรู้คือการก่อตัวของระบบลิมบิกของสมองส่วนหน้าและการก่อตัวของระบบร่างแหของก้านสมอง
ระบบส่งออกของไฮโปทาลามัสยังสามารถแบ่งกลุ่มได้เป็น 3 ทิศทาง:
- ระบบที่ลงสู่การสร้างเรตินูลาร์และไขสันหลัง - ระบบรอบโพรงสมองของใยที่สิ้นสุดที่สมองกลาง (มัดหลังตามยาว) ที่ศูนย์กลางอัตโนมัติของลำต้นส่วนหางและไขสันหลัง และมัดเต้านม-เทกเมนทัล ไปจากบอดีของเต้านมไปยังการสร้างเรตินูลาร์ของสมองกลาง
- เส้นทางสู่ธาลามัสจากมัดเต้านม (มัดเต้านม) ซึ่งเป็นส่วนหนึ่งของระบบลิมบิกแบบทำงานปิด
- เส้นทางสู่ต่อมใต้สมอง - เส้นทางไฮโปทาลามัส-ต่อมใต้สมองจากนิวเคลียสพาราเวนทริคิวลาร์ (10-20% ของใยประสาท) และซูปราออปติก (80-90%) ไปยังกลีบหลังและกลีบกลางบางส่วนของต่อมใต้สมอง เส้นทางทูเบอโรไฮโปไฟเซียลจากนิวเคลียสเวนโตรมีเดียลและอินฟันดิบูลาร์ไปยังอะดีโนไฮโปฟิซิส
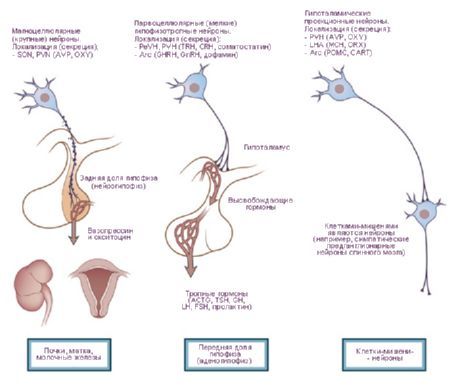
งานของ J. Ranson (1935) และ W. Hess (1930, 1954, 1968) นำเสนอข้อมูลเกี่ยวกับการขยายและหดตัวของรูม่านตา การเพิ่มและลดลงของความดันเลือดแดง การเร่งและชะลอการเต้นของชีพจรเมื่อมีการกระตุ้นไฮโปทาลามัส จากการศึกษาเหล่านี้ พบว่ามีโซนที่ส่งผลต่อระบบซิมพาเทติก (ส่วนหลังของไฮโปทาลามัส) และพาราซิมพาเทติก (ส่วนหน้า) และไฮโปทาลามัสเองถือเป็นศูนย์กลางที่รวมกิจกรรมของอวัยวะและเนื้อเยื่อที่ส่งสัญญาณไปยังระบบอวัยวะภายใน อย่างไรก็ตาม เมื่อการศึกษาเหล่านี้พัฒนาขึ้น ก็พบผลกระทบทางร่างกายจำนวนมากเช่นกัน โดยเฉพาะอย่างยิ่งในระหว่างพฤติกรรมอิสระของสัตว์ [Gellhorn E., 1948] OG Baklavadzhan (1969) สังเกตปฏิกิริยาการกระตุ้นในเปลือกสมองบางส่วนเมื่อมีการกระตุ้นส่วนต่างๆ ของไฮโปทาลามัส ในบางกรณี ทำให้เกิดการตอบสนองต่อการกระตุ้นที่บริเวณเปลือกสมอง ทำให้เกิดศักยภาพโมโนซินแนปส์ของไขสันหลัง ความดันเลือดแดงเพิ่มขึ้น และในบางกรณี มีผลตรงกันข้าม ในกรณีนี้ ปฏิกิริยาตอบสนองแบบ vegetative มีเกณฑ์สูงสุด O. Sager (1962) ค้นพบการยับยั้งของระบบ y และการซิงโครไนซ์ EEG ในระหว่างที่ไฮโปทาลามัสได้รับความร้อน และมีผลตรงกันข้ามในระหว่างที่ร่างกายได้รับความร้อนมากเกินไป แนวคิดที่ว่าไฮโปทาลามัสเป็นส่วนหนึ่งของสมองที่ทำหน้าที่โต้ตอบระหว่างกลไกการควบคุม การบูรณาการกิจกรรมทางร่างกายและทางพืช จากมุมมองนี้ การแบ่งไฮโปทาลามัสไม่ใช่ส่วนซิมพาเทติกและพาราซิมพาเทติก แต่แยกโซนไดนาโมเจนิก (เออร์โกโทรปิกและโทรโฟโทรปิก) ออกจากกันนั้นถูกต้องกว่า การจำแนกประเภทนี้มีลักษณะทางหน้าที่และทางชีววิทยา และสะท้อนถึงการมีส่วนร่วมของไฮโปทาลามัสในการดำเนินการพฤติกรรมแบบองค์รวม เห็นได้ชัดว่าไม่เพียงแต่ระบบพืชเท่านั้น แต่ยังรวมถึงระบบโซมาติกด้วยที่มีส่วนร่วมในการคงสภาพสมดุล โซนเออร์โกโทรปิกและโทรโฟโทรปิกตั้งอยู่ในทุกส่วนของไฮโปทาลามัสและทับซ้อนกันในบางพื้นที่ ในขณะเดียวกัน ก็สามารถระบุโซนของ "สมาธิ" ได้ ดังนั้น ในส่วนหน้า (โซนพรีออปติก) อุปกรณ์โทรโฟโทรปิกจะแสดงได้ชัดเจนยิ่งขึ้น และในส่วนหลัง (บอดีของต่อมน้ำนม) จะแสดงเออร์โกโทรปิก การวิเคราะห์การเชื่อมต่อระหว่างอินพุตและเอาต์พุตหลักของไฮโปทาลามัสกับระบบลิมบิกและเรติคูลัมทำให้เข้าใจบทบาทของไฮโปทาลามัสในการจัดระเบียบรูปแบบพฤติกรรมแบบบูรณาการ ไฮโปทาลามัสมีตำแหน่งพิเศษที่เป็นศูนย์กลางในระบบนี้ เนื่องจากตำแหน่งทางภูมิประเทศอยู่ตรงใจกลางของโครงสร้างเหล่านี้ และเนื่องมาจากลักษณะทางสรีรวิทยา ลักษณะทางสรีรวิทยาถูกกำหนดโดยบทบาทของไฮโปทาลามัสในฐานะส่วนที่ถูกสร้างขึ้นโดยเฉพาะของสมอง โดยไวต่อการเปลี่ยนแปลงของสภาพแวดล้อมภายในร่างกายเป็นพิเศษ โดยตอบสนองต่อการเปลี่ยนแปลงเพียงเล็กน้อยของตัวบ่งชี้ของเหลวในร่างกาย และแสดงพฤติกรรมที่เหมาะสมเพื่อตอบสนองต่อการเปลี่ยนแปลงเหล่านี้บทบาทพิเศษของไฮโปทาลามัสถูกกำหนดล่วงหน้าโดยความใกล้ชิดทางกายวิภาคและการทำงานกับต่อมใต้สมอง นิวเคลียสของไฮโปทาลามัสแบ่งออกเป็นแบบจำเพาะและไม่จำเพาะ กลุ่มแรกประกอบด้วยกลุ่มที่ยื่นออกมาที่ต่อมใต้สมอง ส่วนที่เหลือประกอบด้วยนิวเคลียสอื่น ๆ ซึ่งผลของการกระตุ้นอาจแตกต่างกันไปขึ้นอยู่กับความแรงของแรงกระแทก นิวเคลียสเฉพาะของไฮโปทาลามัสมีผลชัดเจนและแตกต่างจากกลุ่มอื่น ๆ ของสมองในความสามารถในการสร้างเส้นประสาทคริเนีย ได้แก่ นิวเคลียสเหนือออปติก พาราเวนทริคิวลาร์ และพาร์โวเซลลูลาร์ของตุ่มสีเทา ได้มีการพิสูจน์แล้วว่าฮอร์โมนต่อต้านการขับปัสสาวะ (ADH) ก่อตัวในนิวเคลียสเหนือออปติกและพาราเวนทริคิวลาร์ ซึ่งเคลื่อนลงมาตามแอกซอนของทางเดินไฮโปทาลามัส-ต่อมใต้สมองไปยังกลีบหลังของต่อมใต้สมอง ต่อมามีการแสดงให้เห็นว่าปัจจัยการปลดปล่อยเกิดขึ้นในเซลล์ประสาทของไฮโปทาลามัส ซึ่งเมื่อเข้าสู่ต่อมใต้สมองส่วนหน้า จะควบคุมการหลั่งฮอร์โมนสามชนิด ได้แก่ อะดรีโนคอร์ติโคโทรปิก (ACTH), ลูทีไนซิง (LH), ฟอลลิเคิลสติมูเลติ้ง (FSH) และไทรอยด์สติมูเลติ้ง (TSH) โซนของการก่อตัวของปัจจัยที่มีผลต่อ ACTH และ TSH คือ นิวเคลียสของส่วนหน้าของเนินนูนกลางและบริเวณพรีออปติก และสำหรับ GTG คือส่วนหลังของตุ่มสีเทา ได้มีการพิสูจน์แล้วว่ามัดไฮโปทาลามัส-ต่อมใต้สมองในมนุษย์มีเส้นใยประสาทประมาณ 1 ล้านเส้น
ไม่ต้องสงสัยเลยว่าส่วนอื่นๆ ของสมอง (โครงสร้างส่วนกลาง-ฐานของบริเวณขมับ โครงร่างเรตินูลาร์ของก้านสมอง) ยังมีส่วนร่วมในการควบคุมระบบประสาทต่อมไร้ท่อด้วย อย่างไรก็ตาม อุปกรณ์ที่เฉพาะเจาะจงที่สุดคือไฮโปทาลามัส ซึ่งรวมถึงต่อมไร้ท่อในระบบปฏิกิริยารวมของร่างกาย โดยเฉพาะอย่างยิ่งปฏิกิริยาที่มีลักษณะเครียด ระบบโทรโฟและเออร์โกโทรปิกมีไม่เพียงแต่ระบบซิมพาเทติกและพาราซิมพาเทติกรอบนอกเท่านั้นที่จะช่วยให้เกิดการทำงาน แต่ยังมีอุปกรณ์ทางระบบประสาทฮอร์โมนเฉพาะอีกด้วย ระบบไฮโปทาลามัส-ต่อมใต้สมอง ซึ่งทำงานบนหลักการของการตอบรับ มีหน้าที่ในการควบคุมตนเองเป็นส่วนใหญ่ กิจกรรมของการก่อตัวของปัจจัยที่นำไปใช้ยังถูกกำหนดโดยระดับของฮอร์โมนในเลือดรอบนอกอีกด้วย
ดังนั้น ไฮโปทาลามัสจึงเป็นส่วนประกอบสำคัญของระบบลิมบิกและระบบเรตินูลาร์ของสมอง แต่เมื่อรวมอยู่ในระบบเหล่านี้ ไฮโปทาลามัสยังคงรักษา "ข้อมูลเข้า" เฉพาะไว้ในรูปแบบของความไวพิเศษต่อการเปลี่ยนแปลงในสภาพแวดล้อมภายใน เช่นเดียวกับ "ข้อมูลออก" เฉพาะผ่านระบบไฮโปทาลามัส-ต่อมใต้สมอง การเชื่อมต่อพาราเวนทริคิวลาร์ไปยังการก่อตัวทางพืชที่อยู่ด้านล่าง รวมทั้งผ่านทาลามัสและการสร้างเรตินูลาร์ของก้านสมองไปยังคอร์เทกซ์และไขสันหลัง
 [
[ สิ่งที่ต้องตรวจสอบ?