ผู้เชี่ยวชาญทางการแพทย์ของบทความ

สิ่งตีพิมพ์ใหม่
ไวรัสเอชไอวี (human immunodeficiency virus)
ตรวจสอบล่าสุด: 23.04.2024

เนื้อหา iLive ทั้งหมดได้รับการตรวจสอบทางการแพทย์หรือตรวจสอบข้อเท็จจริงเพื่อให้แน่ใจว่ามีความถูกต้องตามจริงมากที่สุดเท่าที่จะเป็นไปได้
เรามีแนวทางการจัดหาที่เข้มงวดและมีการเชื่อมโยงไปยังเว็บไซต์สื่อที่มีชื่อเสียงสถาบันการวิจัยทางวิชาการและเมื่อใดก็ตามที่เป็นไปได้ โปรดทราบว่าตัวเลขในวงเล็บ ([1], [2], ฯลฯ ) เป็นลิงก์ที่คลิกได้เพื่อการศึกษาเหล่านี้
หากคุณรู้สึกว่าเนื้อหาใด ๆ ของเราไม่ถูกต้องล้าสมัยหรือมีข้อสงสัยอื่น ๆ โปรดเลือกแล้วกด Ctrl + Enter
ได้รับโรคภูมิคุ้มกันบกพร่องที่ได้รับมาเป็นโรคเฉพาะในปีพ. ศ. 2524 ในสหรัฐอเมริกาเมื่อมีคนเป็นโรคร้ายแรงจำนวนมากเกิดจากเชื้อจุลินทรีย์ที่ไม่ก่อให้เกิดโรคหรือก่อให้เกิดโรคเล็กน้อยสำหรับคนที่มีสุขภาพ การตรวจสอบสถานะภูมิคุ้มกันของผู้ป่วยพบว่ามีจำนวน lymphocytes ลดลงมากและผู้ช่วย T โดยเฉพาะอย่างยิ่ง ภาวะนี้เรียกว่าโรคเอดส์ (โรคภูมิคุ้มกันบกพร่องที่ได้มาจากภาษาอังกฤษ - โรคภูมิคุ้มกันบกพร่องที่ได้มาหรือเอดส์) วิธีการติดเชื้อ (ติดต่อทางเพศผ่านผลิตภัณฑ์เลือดและเลือด) ชี้ให้เห็นลักษณะการติดเชื้อของโรค
สาเหตุของโรคเอดส์ถูกค้นพบในปี 2526 โดยชาวฝรั่งเศส L. Montagnier ผู้ซึ่งตั้งชื่อเขาว่า LAV Lymphoadenopathy Associated Virus) เนื่องจากเขาตรวจพบผู้ป่วยที่มี lymphadenopathy; (American Human T-lymphotropic Virus III) ซึ่งก่อนหน้านี้เขาได้ค้นพบไวรัส lymphotropic I และ II
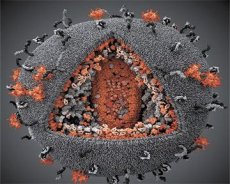
การเปรียบเทียบสมบัติของไวรัส LAV และ HTLV-III มีความเป็นตัวตนดังนั้นในปี พ.ศ. 2529 ไวรัสจึงถูกเรียกว่าเอชไอวี (Human Immunodeficiency Virus หรือ HIV) เพื่อหลีกเลี่ยงความสับสน เอชไอวีทรงกลมมีเส้นผ่านศูนย์กลาง 110 นาโนเมตร ซองจดหมายของไวรัสมีรูปทรงหลายเหลี่ยมประกอบด้วย 12 แฉกและรูป 20 เหลี่ยม โมเลกุลของโปรตีน glycosylated gpl20 ตั้งอยู่ตรงกลางและมุมของแต่ละเหลี่ยม (จำนวน 120 หมายถึงน้ำหนักโมเลกุลของโปรตีนใน kilodaltons) มีทั้งหมด 72 gpl20 โมเลกุลตั้งอยู่บนพื้นผิวของ virion ซึ่งแต่ละอันเกี่ยวข้องกับโปรตีน intramembrane gp41 โปรตีนเหล่านี้ร่วมกับชั้นไขมันสองชั้นสร้าง supercapsid (membrane) ของ virion
โปรตีน gpl20 และ gp41 เกิดขึ้นจากการตัดโปรตีนโปรตีเอสของโปรตีนสารตั้งต้น Env. โปรตีน gp41 สร้าง "แกน" ของกระดูกสันหลังที่เชื่อมโยงกันโดยโดเมน cytoplasmic ด้วยเมทริกซ์โปรตีน p17MA ทันทีใต้เมมเบรน โมเลกุล p17, ปฏิสัมพันธ์กับการเจริญเติบโตของ virion, รูปแบบ icosahedron เปลือกต้นแบบ
ในภาคกลางของ virion, โปรตีน p24 สร้างรูปทรงกรวย ส่วนที่แคบลงของ capsid ที่มีส่วนร่วมของโปรตีน pb เชื่อมต่อกับซองจดหมายของ virion ภายใน capsid มีโมเลกุลสองตัวที่เหมือนกันของ RNA ของยีนไวรัส พวกเขาถูกผูกไว้ที่ปลาย 5 'ของพวกเขาไปยังโปรตีน nucleocapsid p7NC โปรตีนชนิดนี้เป็นที่น่าสนใจในการที่จะมีสองกรดอะมิโนตกค้าง (บรรทัดฐาน) cysteine ที่อุดมด้วยและฮิสติดีนและอะตอมที่มีธาตุสังกะสี - พวกเขาจะเรียกว่า "สังกะสีนิ้ว" ขณะที่พวกเขาจับโมเลกุลของ RNA จีโนมสำหรับการรวมตัวเข้ามาในรูปแบบที่ virions แคปซูลยังประกอบด้วยเอนไซม์สามตัว Reversase (RT) หรือ pol complex ประกอบด้วย transcriptase แบบย้อนกลับ, RNA-ase H และ DNA polymerase polymerase revertase มีอยู่ในรูปแบบ p66 / p51 heterodimer Protease (PR) - pI เริ่มต้นและตระหนักถึงกระบวนการของการเจริญเติบโตของเชื้อรา การรวมเข้าด้วยกัน (IN) - p31 หรือ endonuclease ช่วยให้แน่ใจได้ว่าการรวม proviral DNA เข้ากับจีโนมของเซลล์เจ้าบ้าน capsid ยังมีโมเลกุลของ RNA เมล็ด (tRNAl "3)
ยีน RNA ในเซลล์ถูกแปลงโดย reverse transcriptase เป็น DNA genome (DNA-provirus) ประกอบด้วย 9283 nucleotide pair จะถูก จำกัด ไว้ที่ด้านซ้ายและด้านขวาโดยการเรียกซ้ำแบบ long end หรือ LTR (การทำซ้ำแบบ terminal ระยะยาวของภาษาอังกฤษ): S'-LTR - ซ้ายและ Z'-LTR - ทางด้านขวา LTR มี 638 nucleotide pair
จีโนมของเอชไอวีประกอบด้วยยีน 9 ตัวซึ่งบางส่วนมีการซ้อนทับกันอยู่ (มีกรอบการอ่านหนังสืออยู่หลายรูปแบบ) และมีโครงสร้างของ exonine พวกเขาควบคุมการสังเคราะห์โปรตีนโครงสร้าง 9 และ 6 ตัว
ค่า LTR ของจีโนมไวรัสคือว่ามีองค์ประกอบด้านกฎข้อบังคับต่อไปนี้ซึ่งควบคุมการทำงานของไวรัส:
- สัญญาณถอดรหัส (ภูมิภาคโปรโมเตอร์);
- สัญญาณสำหรับการเติมโพลี - เอ
- จับสัญญาณ;
- การรวมสัญญาณ
- สัญญาณควบคุมเชิงบวก (TAR for TAT protein);
- องค์ประกอบของกฎเกณฑ์เชิงลบ (NRE for NEF protein);
- ตำแหน่งที่แนบมาของเมล็ดอาร์เอ็นเอ (tRNA ™ 3) สำหรับการสังเคราะห์ดีเอ็นเอลบโซ่ที่ปลาย 3 '; สัญญาณที่ 5'-end ของ LTR ซึ่งทำหน้าที่เป็นไพรเมอร์สำหรับการสังเคราะห์ดีเอ็นเอบวกเส้นใย
นอกจากนี้ LTR มีองค์ประกอบที่เกี่ยวข้องกับการควบคุมการ splicing ของ mRNA ซึ่งบรรจุโมเลกุล vRNA ไว้ใน capsid (องค์ประกอบของ Psi) ในที่สุดเมื่อถ่ายทอดโครโมโซมใน mRNAs ยาว ๆ จะมีการสร้างสัญญาณสองตัวสำหรับโปรตีน REV ซึ่งจะสลับการสังเคราะห์โปรตีน: CAR สำหรับโปรตีนควบคุมและ CRS สำหรับโปรตีนโครงสร้าง ถ้าโปรตีน REV เชื่อมโยงกับ CAR จะมีการสังเคราะห์โปรตีนโครงสร้าง ถ้าไม่มีอยู่จะมีการสังเคราะห์โปรตีนตามข้อบังคับเท่านั้น
ในกฎระเบียบของจีโนมของไวรัสยีนที่ควบคุมต่อไปนี้และโปรตีนของพวกเขามีบทบาทสำคัญอย่างยิ่ง:
- โปรตีน TAT ที่มีประสิทธิภาพในการควบคุมการสืบพันธุ์ของไวรัสและทำหน้าที่ผ่านเว็บไซต์ TAR ที่มีการควบคุม
- โปรตีน NEV และ VPU ดำเนินการควบคุมการสืบพันธุ์ในเชิงลบผ่านพื้นที่ NRE;
- โปรตีน REV, การดำเนินการควบคุมบวกลบ โปรตีน REV ควบคุมการทำงานของยีนส์ปิดปาก pol, env และดำเนินการควบคุมเชิงลบของ splicing
ดังนั้นการทำสำเนาของเอชไอวีอยู่ภายใต้การควบคุมสาม - บวก, ลบและบวกลบ
โปรตีน VIF กำหนดความสามารถในการติดเชื้อของไวรัสที่สังเคราะห์ขึ้นใหม่ มันผูกพันกับโปรตีน capsid p24 และมีอยู่ใน virion ในจำนวน 60 โมเลกุล โปรตีน NEF มีอยู่ใน virion โดยมีโมเลกุลจำนวนน้อย (5-10) ซึ่งอาจเชื่อมต่อกับซองจดหมาย
โปรตีน VPR ยับยั้งวงจรมือถือที่ระยะ G2 มีส่วนร่วมในคอมเพล็กซ์ขนส่ง preintegratsionnyh ในนิวเคลียสของเซลล์และเปิดใช้งานยีนไวรัสและเซลล์บางอย่างเพิ่มประสิทธิภาพของการจำลองแบบของไวรัสใน monocytes และ macrophages ไม่พบตำแหน่งของโปรตีน VPR, TAT, REV, VPU ใน virion
นอกเหนือไปจากโปรตีนของตัวเององค์ประกอบของเมมเบรน virion อาจรวมถึงโปรตีนบางชนิดของเซลล์เจ้าบ้าน โปรตีน VPU และ VPR มีส่วนเกี่ยวข้องกับการควบคุมการสืบพันธุ์ของไวรัส
ตัวแปรของแอนติเจนของไวรัสภูมิคุ้มกันบกพร่องของมนุษย์ (HIV)
ไวรัสภูมิคุ้มกันบกพร่องของมนุษย์ (HIV) มีความแตกต่างกันมาก แม้แต่จากเชื้อโรคของผู้ป่วยรายใดรายหนึ่งสามารถแยกเชื้อไวรัสที่มีความแตกต่างกันอย่างมีนัยสำคัญทางชีวเคมี ความผันแปรดังกล่าวได้รับการส่งเสริมโดยการทำลายเซลล์ CD4 + อย่างเข้มข้นและการตอบสนองของแอนติบอดีที่มีประสิทธิภาพต่อการติดเชื้อเอชไอวี ผู้ป่วยจากแอฟริกาตะวันตกมีรูปแบบเอชไอวีใหม่ทางชีวภาพใกล้กับเอชไอวี -1 แต่แตกต่างจากภูมิคุ้มกันเชื้อ HIV-2 ความคล้ายคลึงกันของโครงสร้างหลักของจีโนมของไวรัสเหล่านี้คือ 42% DNA-provirus HIV-2 มี 9671 bp และ LTR-854 bp HIV-2 ถูกแยกจากกันในภูมิภาคอื่น ๆ ของโลก ไม่มีภูมิคุ้มกันข้ามระหว่างเอชไอวี -1 และเอชไอวี -2 รูปที่ 2 ของ HIV-1 เป็นที่รู้จักกันดี: O (Outlier) และ M (Major) ส่วนหลังแบ่งเป็น 10 subtypes (AJ) ในรัสเซียมีอนุภาคย่อย 8 ตัว (AH)
 [
[กลไกการปฏิสัมพันธ์ของเอชไอวีกับเซลล์
เมื่อเจาะเข้าไปในร่างกายไวรัสจะโจมตีเซลล์แรกที่มีตัวรับ CD4 ที่เฉพาะเจาะจง ตัวรับนี้มีผู้ช่วย T จำนวนมากในจำนวนที่น้อยกว่า - macrophages และ monocytes โดยเฉพาะอย่างยิ่งผู้ช่วย T ที่ไวต่อไวรัส
ไวรัสภูมิคุ้มกันบกพร่องของมนุษย์ (HIV) ตระหนักถึงผู้รับ CD4 ด้วยโปรตีน gpl20 กระบวนการปฏิสัมพันธ์ของเอชไอวีกับเซลล์ดำเนินไปตามรูปแบบดังต่อไปนี้: การดูดซับตัวรับ -> ที่มีชายแดนล้อมรอบ -> ถุงที่มีรอยแต้ม -> lysosome เมมเบรนของ virion ผสานกับเมมเบรน lysosome และ nucleocapsid ซึ่งเป็นอิสระจาก supercapsid เข้าไปใน cytoplasm; ระหว่างทางไปนิวเคลียสจะถูกทำลายและจะมีการปลดปล่อยอาร์เอ็นเอจีโนมิกและองค์ประกอบหลักที่เกี่ยวข้อง จากนั้นการถอดรหัสย้อนกลับจะสังเคราะห์ดีเอ็นเอของ DNA บนไวรัส RNA จากนั้น RNA-ase H จะทำลาย virion RNA และ DNA polymerase ของไวรัสจะสังเคราะห์เส้นใยบวกของ DNA ในตอนท้ายของ DNA-provirus จะสร้าง 5'-LTR และ 3'-LTR DNA provirus สามารถอยู่ในนิวเคลียสในขณะที่อยู่ในรูปแบบที่ไม่ใช้งาน แต่ไม่ช้าก็เร็วมันรวมกับความช่วยเหลือของ integrase ในโครโมโซมของเซลล์เป้าหมาย ในนั้น provirus อยู่ในสถานะที่ไม่ใช้งานจนกว่า T-lymphocyte นี้จะถูกกระตุ้นโดยแอนติเจนเชื้อจุลินทรีย์หรือเซลล์ภูมิคุ้มกันอื่น ๆ การเปิดใช้งานการถอดความ DNA ของโทรศัพท์มือถือถูกควบคุมโดยปัจจัยนิวเคลียร์พิเศษ (NF-kB) เป็นโปรตีนที่มีผลผูกพัน DNA และผลิตในปริมาณมากระหว่างการกระตุ้นและการขยายตัวของ T-lymphocytes และ monocytes โปรตีนนี้เชื่อมโยงกับลำดับเฉพาะของดีเอ็นเอของเซลล์และลำดับเดียวกันของ LTR DNA-provirus และทำให้เกิดการถอดรหัสของดีเอ็นเอของเซลล์และ DNA provirus โดยการกระตุ้นการถอดความของ DNA provirus เขาเปลี่ยนไวรัสจากรัฐที่ไม่ได้ใช้งานไปสู่การใช้งานและดังนั้นการติดเชื้อแบบถาวรลงในการผลิตอย่างใดอย่างหนึ่ง พัก provirus ในรัฐที่ไม่ใช้งานสามารถใช้เวลานานมาก การเปิดใช้งานของไวรัสเป็นช่วงเวลาที่สำคัญในการปฏิสัมพันธ์กับเซลล์
ตั้งแต่เวลาที่เชื้อไวรัสเข้าสู่เซลล์ระยะเวลาของการติดเชื้อเอชไอวีจะเริ่มขึ้นซึ่งเป็นไวรัสที่มีอายุ 10 ปีหรือมากกว่านั้น และตั้งแต่การกระตุ้นของไวรัสจะเริ่มเป็นโรค - โรคเอดส์ ด้วยความช่วยเหลือของยีนการกำกับดูแลและผลิตภัณฑ์ของตนไวรัสจึงเริ่มทวีความรุนแรงมากขึ้น TAT-protein สามารถเพิ่มอัตราการสืบพันธุ์ของไวรัสได้ 1000 เท่า การถอดความของไวรัสมีความซับซ้อน ประกอบด้วยการสร้างทั้ง mRNAs ความยาวเต็มและ subgenomic, splicing ของ mRNA และการสังเคราะห์โปรตีนโครงสร้างและกฎระเบียบเพิ่มเติม
การสังเคราะห์โปรตีนโครงสร้างมีดังนี้ ครั้งแรกจะมีการสังเคราะห์โปรตีน Pr55Gag ซึ่งเป็นโปรตีนที่มีมวล 55 kD มันมีสี่โดเมนหลัก: เมทริกซ์ (MA) capsid (CA) นิวคลีโอ (NC) และ PE โดเมนซึ่งส่งผลให้เกิดการตัด Pr55Gag ไวรัสโปรติเอส (มัน samovyrezaetsya จากโปรตีนอื่นปูชนียบุคคล - Gag-Pol) จะเกิดขึ้นตามลำดับโปรตีนโครงสร้าง P17 , p24, p7 และ pb การก่อตัว Polyprotein Pr55Gag เป็นเงื่อนไขหลักในการก่อตัวของอนุภาคไวรัส มันเป็นโปรตีนที่กำหนดโปรแกรมของ morphogenesis ของ virion นี้ ซึ่งจะรวมถึงขั้นตอนของการขนส่ง polyprotein ลำดับปิดปากเพื่อเยื่อหุ้มพลาสม่าและการมีปฏิสัมพันธ์กับปฏิกริยาระหว่างโปรตีนในการก่อตัวของอนุภาคไวรัสและรุ่นของตน Pr55Gag สังเคราะห์ในโพลีเบอโรเมียฟรี โมเลกุลโปรตีนจะถูกส่งไปยังเมมเบรนที่พวกเขายึดตัวเองกับแพทช์ไม่ชอบน้ำของพวกเขา บทบาทหลักในการสร้างรูปแบบดั้งเดิมของโปรตีน Gag ถูกเล่นโดยโดเมน CA สวิทช์ NC-โดเมนให้ (โดยวิธีการของ "นิ้วสังกะสี") 2 โมเลกุลของ RNA จีโนมเข้าไปในอนุภาคไวรัสที่ก่อตัวขึ้น โมเลกุลโพรพิลีนเป็นครั้งแรก dimerized เนื่องจากปฏิสัมพันธ์ของโดเมนเมทริกซ์ จากนั้น dimers รวมกันเป็น hexameric (6 หน่วย) คอมเพล็กซ์โดยมีปฏิสัมพันธ์โดเมนแคลิฟอร์เนียและอร์ทแคโรไลนา สุดท้าย hexamers เชื่อมต่อพื้นผิวด้านประกอบ virions อ่อนรูปทรงกลมภายในซึ่งมีจีโนมของไวรัส RNA จับ NC-โดเมน
โปรตีนเป็นสารตั้งต้นอีก Prl60Gag-Pol (โปรตีนที่มีม. เอ็ม 160 กิโลดาลตัน) ถูกสังเคราะห์เป็นผลมาจากการขยับกรอบการอ่านของไรโบโซมในระหว่างการแปล z 'ปลายทางของยีนปิดปากลงไปในพื้นที่ทันทีก่อนที่จะเข้ารหัสโปรตีน RB ภูมิภาค โพรพิลีน Gag-Pol นี้ประกอบด้วยลำดับโปรตีนที่ไม่สมบูรณ์ (1 - 423 amino acids) ที่ไม่สมบูรณ์และลำดับของ Pol ซึ่งรวมถึงโดเมน PR, RT และ IN โมเลกุลของ polyprotein Gag-Pol ยังถูกสังเคราะห์ขึ้นใน polyribosomes ฟรีและนำไปพลาสม่าเมมเบรน Polyprotein Prl60Gagpol มีตำแหน่งทั้งหมดของปฏิสัมพันธ์ระหว่างโมเลกุลที่อยู่ในโพรพิลีน Gag และบริเวณที่มีเยื่อหุ้มเซลล์ ดังนั้นโมเลกุลของ polyprotein ฟิวส์ Gag-Pol ด้วยเมมเบรนและร่วมกับ Gag โมเลกุล ได้แก่ virions ขึ้นรูปซึ่งอาจส่งผลในการใช้งานโปรติเอสและการเจริญเติบโต virion กระบวนการเริ่มต้น เอชไอวี -1 โปรติเอสที่ใช้งานสูงเป็นเพียง dimer จึง samovyrezaniya ของ Prl60Gag-Pol ต้อง dimerization ของโมเลกุลเหล่านี้ การสุกของเชื้อไวรัสคือการที่โปรตีนที่ปล่อยออกมาได้รับการปลดปล่อย prl60Gag-Pol และ Gag55 เข้าไปในบริเวณที่เป็นที่รู้จัก โปรตีน p17, p24, p7, p6, revertase, integrase จะเกิดขึ้นและความสัมพันธ์ของพวกเขาในโครงสร้างของไวรัสจะเกิดขึ้น
Env โปรตีนสังเคราะห์ในไรโบโซมที่เกี่ยวข้องกับเยื่อของร่างแหเอนโดพลาซึมที่ glycosylated และจากนั้นก็จะถูกตัดโปรติเอสมือถือเพื่อ gp120 และ gp41 และจะถูกส่งให้กับเซลล์ผิว ในกรณีนี้ gp41 แทรกซึมเมมเบรนและเชื่อมโยงกับโดเมนเมทริกซ์ของโมเลกุลโปรตีน Gag ที่เกี่ยวข้องกับผิวด้านในของเมมเบรน ความสัมพันธ์นี้ยังคงมีอยู่ในผู้ใหญ่
ดังนั้นการชุมนุมของอนุภาคไวรัสคือการรวมตัวของโปรตีนสารตั้งต้นและโมเลกุล RNA ที่เกี่ยวข้องบนเยื่อหุ้มเซลล์โฮสต์การก่อตัวของ virions ที่ยังไม่บรรลุนิติภาวะและการเปิดตัวของพวกเขาโดยการแตกหน่อจากเซลล์ผิว เมื่อเกิดไวรัส virion ล้อมรอบตัวเองด้วยเมมเบรนของเซลล์ซึ่งมีโมเลกุล gp41 และ gp120 ฝังอยู่ ในช่วงออกดอกหรืออาจจะเป็นหลังจากการเปิดตัวของ virion การเจริญเติบโตที่เกิดขึ้นซึ่งจะดำเนินการโดยใช้เอนไซม์โปรติเอไวรัสถูกตัดโปรตีนโปรตีน Pr55Gag สารตั้งต้นและไวรัส Prl60Gag-Pol ผู้ใหญ่โปรตีนและความสัมพันธ์ของพวกเขาเพื่อคอมเพล็กซ์โครงสร้างที่เฉพาะเจาะจง บทบาทสำคัญในกระบวนการของไวรัส morphogenesis เล่นโดยโพรพิลีนสารตั้งต้น Pr55Gag ซึ่งจัดและประกอบ virion ยังไม่บรรลุนิติภาวะ; กระบวนการของการเจริญเติบโตเสร็จสมบูรณ์โดยโปรโตคอลไวรัสที่เฉพาะเจาะจง
สาเหตุของภาวะภูมิคุ้มกันบกพร่อง
หนึ่งในสาเหตุหลักของการติดเชื้อเอชไอวีในผู้ป่วยด้วยโรคภูมิคุ้มกันบกพร่องคือการเสียสละมวลของผู้ช่วยที เกิดขึ้นเนื่องจากเหตุการณ์ต่อไปนี้ ประการแรกไวรัส T-helper ที่ติดเชื้อไวรัสตายเนื่องจาก apoptosis เป็นที่เชื่อกันว่าในผู้ป่วยโรคเอดส์การจำลองแบบของไวรัส apoptosis และการลดจำนวนผู้ช่วย T ที่เกี่ยวข้อง ประการที่สอง T เซลล์นักฆ่ารับรู้และทำลาย T-เซลล์ที่ติดเชื้อไวรัสหรือแบริ่งโมเลกุลดูดซับ gpl20 เช่นเดียวกับการติดเชื้อไวรัสและเซลล์ T-ผู้ช่วยที่ติดเชื้อไวรัสซึ่งรูปแบบ symplasts (syncytia) ประกอบด้วยหลายสิบของเซลล์ (ส่วนหนึ่งของ พวกเขาตายอันเป็นผลมาจากการเพิ่มจำนวนของไวรัสในพวกเขา) เนื่องจากการทำลายของจำนวนมากของเซลล์ T-ผู้ช่วยที่เกิดขึ้นการแสดงออกของการลดลงของเมมเบรนรับในเซลล์เม็ดเลือดขาว B เพื่อ IL-2, รบกวนการสังเคราะห์ interleukins ต่างๆ (ปัจจัยการเจริญเติบโตและความแตกต่างของ B-lymphocytes. - IL-4, IL-5, IL-6, และอื่น ๆ ) เป็นผลมาจากการที่การทำงานของระบบ T-killer ถูกละเมิด การปราบปรามการทำงานของระบบเสริมและระบบ macrophage เกิดขึ้น macrophages ที่ติดไวรัสและ monocytes ไม่ตายเป็นเวลานาน แต่พวกเขาไม่สามารถเอาไวรัสออกจากร่างกายได้ ในที่สุดเนื่องจากความคล้ายคลึงกันโครงสร้างและแอนติเจนกับตัวรับ gpl20 บางเซลล์เยื่อบุผิวของสิ่งมีชีวิต (รวม trophoblasts รับเป็นสื่อกลางในการส่งของการปลูกถ่ายเอชไอวี) ถูกสังเคราะห์แอนติบอดี antiretseptornyh กับคลื่นความถี่กว้างของการกระทำ แอนติบอดีดังกล่าวสามารถป้องกันตัวรับเซลล์ต่างๆและทำให้เกิดความซับซ้อนของโรคที่มีความผิดปกติของภูมิต้านตนเองได้ ผลของการติดเชื้อเอชไอวีคือความพ่ายแพ้ของส่วนที่สำคัญทั้งหมดของระบบภูมิคุ้มกัน ผู้ป่วยดังกล่าวสามารถป้องกันตัวจุลินทรีย์ได้หลากหลายชนิด นี้นำไปสู่การพัฒนาของการติดเชื้อฉวยโอกาสและโรคประสาท สำหรับผู้ป่วยที่ติดเชื้อเอชไอวีโรคมะเร็งอย่างน้อย 3 ชนิดมีความเสี่ยงเพิ่มขึ้น ได้แก่ Kaposi's sarcoma; มะเร็ง (รวมถึงมะเร็งผิวหนัง); มะเร็งต่อมน้ำเหลือง B-cell ที่เกิดจากความเสื่อมของมะเร็งต่อมน้ำเหลืองของ B-lymphocytes อย่างไรก็ตามเอชไอวีไม่เพียง แต่มีเม็ดเลือดขาว แต่ยังเป็นโรคประสาท มันแทรกซึมในระบบประสาทส่วนกลางเซลล์ (astrocytes) ทั้งโดยการนำสารเข้าสู่เซลล์แบบใช้ตัวรับและเซลล์ทำลายของ astrocytes กับ lymphoblasts ที่ติดเชื้อไวรัส เมื่อไวรัสมีปฏิสัมพันธ์กับ astrocytes symplasts จะเกิดขึ้นซึ่งช่วยในการแพร่กระจายเชื้อโรคผ่านช่องทางระหว่างเซลล์ ในขนาดใหญ่และ monocytes ไวรัสอาจยังคงมีอยู่เป็นเวลานานเพื่อให้พวกเขาทำหน้าที่เป็นอ่างเก็บน้ำและผู้จัดจำหน่ายในร่างกาย, ความสามารถในการเจาะเข้าไปในเนื้อเยื่อ macrophages ที่ติดเชื้อมีบทบาทสำคัญในการอพยพของเอชไอวีในระบบประสาทส่วนกลางและความพ่ายแพ้ ใน 10% ของผู้ป่วยอาการทางคลินิกหลักเกี่ยวข้องกับความเสียหายของระบบประสาทส่วนกลางและการแสดงออกเป็นภาวะสมองเสื่อม (dementia) ดังนั้นสำหรับผู้ที่ติดเชื้อเอชไอวีมี 3 กลุ่มคือโรคติดเชื้อฉวยโอกาสโรคเนื้องอกและความเสียหายของระบบประสาทส่วนกลาง
ระบาดวิทยาของการติดเชื้อเอชไอวี
แหล่งที่มาของการติดเชื้อเอชไอวีเป็นเพียงผู้ป่วยหรือผู้ให้บริการไวรัสเท่านั้น ไวรัสภูมิคุ้มกันบกพร่องของมนุษย์ (HIV) พบได้ในเลือดน้ำอสุจิปากมดลูก ในมารดาที่เลี้ยงดู - ในนมแม่ การติดเชื้อเกิดขึ้นทางเพศผ่านทางเลือดและยาเสพติดรวมทั้งจากมารดาถึงเด็กก่อนคลอดระหว่างและหลังคลอด กรณีที่มีการติดเชื้อไวรัสโดยอาหารเครื่องดื่มและแมลงกัดไม่ทราบ
การติดยาเสพติดก่อให้เกิดการแพร่กระจายของโรคเอดส์ การติดเชื้อเอชไอวีจะเพิ่มขึ้นทุกปี อ้างอิงจาก WHO ในช่วงปีพ. ศ. 2523 ถึงปี พ.ศ. 2543 มีผู้ติดเชื้อเอดส์จำนวน 58 ล้านคน เฉพาะในปีพ. ศ. 2543 มีผู้ติดเชื้อ 5.3 ล้านคนทั่วโลกและ 3 ล้านคนเสียชีวิตจากโรคเอดส์ ในรัสเซียเมื่อวันที่ 1 มกราคม 2547 มีผู้ลงทะเบียนเชื้อเอชไอวี 264,000 ราย ครึ่งหนึ่งของผู้ติดเชื้อเอชไอวีเสียชีวิตภายใน 11-12 ปีของการติดเชื้อ ในช่วงต้นปี 2547 ประชาชนชาวรัสเซีย 100,000 รายได้รับการวินิจฉัยว่าติดเชื้อเอชไอวีประมาณ 180 ราย คาดว่าในระดับนี้จะมีผู้ติดเชื้อเอชไอวีจำนวนประมาณ 2.5-3 ล้านคนในปีพ. ศ. 2555 ความซับซ้อนของการต่อสู้กับการติดเชื้อเอชไอวีขึ้นอยู่กับหลายสาเหตุ: ประการแรกไม่มีวิธีการรักษาและการป้องกันที่มีประสิทธิภาพ ประการที่สองระยะฟักตัวในการติดเชื้อเอชไอวีจะเกิน 10 ปี ระยะเวลาขึ้นอยู่กับช่วงเวลาของการกระตุ้นของ T-lymphocyte และ DNA provirus ที่มีอยู่ในโครโมโซมของมัน ยังไม่เป็นที่แน่ชัดว่าไวรัสแต่ละตัวติดเชื้อเอดส์จะถึงวาระหรือมีแนวโน้มที่จะมีไวรัสระยะยาวโดยไม่มีโรค (ซึ่งดูเหมือนว่าไม่น่าจะเป็นไปได้) สุดท้ายมีไวรัสภูมิคุ้มกันบกพร่องของมนุษย์จำนวนมาก (HIV-1, HIV-2), ความแตกต่างระหว่างแอนติบอดีซึ่งทำให้เกิดภูมิคุ้มกันข้าม การตรวจหาไวรัสโรคภูมิคุ้มกันบกพร่องของลิง (SIV) ทำให้เกิดการติดเชื้อเอชไอวี SIO สำหรับการจัดระเบียบจีโนมคล้ายคลึงกับเอชไอวี แต่มันแตกต่างกันอย่างมีนัยสำคัญในลำดับนิวคลีโอไทด์ ซีรั่มเอชไอวี -2 อยู่ในตำแหน่งกลางระหว่าง HIV-1 และ SIV และลำดับของ nucleotide ก็ใกล้เคียงกับ SIV ในเรื่องนี้ VM Zhdanov ชี้ให้เห็นว่าไวรัส HIV - 1, HIV - 2 และ SIV มาจากบรรพบุรุษร่วมกัน มีความเป็นไปได้ที่อาร์กัลโล (R. Gallo) คนหนึ่งของ SIVs ได้เข้าสู่ร่างกายมนุษย์ซึ่งเป็นที่มาของการกลายพันธุ์ซึ่งทำให้เกิด HIV-1, HIV-2 และรูปแบบอื่น ๆ
อาการของการติดเชื้อเอชไอวี
ไวรัสของภูมิคุ้มกันบกพร่องของมนุษย์มีคุณลักษณะบางอย่างซึ่งเป็นสาเหตุของโรคส่วนใหญ่ ไวรัสมีอัตราการสืบพันธุ์สูงมากโดยพิจารณาจากองค์ประกอบด้านกฎระเบียบ (5,000 virion จะถูกสังเคราะห์ภายใน 5 นาทีในระยะที่ใช้งาน) เนื่องจากมีโปรตีน fusion (gp41) ไวรัสทำให้เกิดโครงสร้าง syncytial ที่กว้างขวางขึ้นเนื่องจากการหลอมรวมของ T-aiders ที่ติดเชื้อและไม่ติดเชื้อซึ่งส่งผลให้เกิดการเสียชีวิตจำนวนมาก โมเลกุลจีพีเอสโมเลกุลโมเลกุลของโมเลกุลขนาดใหญ่สามารถไหลเวียนได้อย่างอิสระในเลือดและผูกติดกับตัวรับ T ผู้ที่ไม่ติดเชื้อซึ่งเป็นผลให้พวกเขาได้รับการยอมรับและถูกทำลายด้วย T-killers ไวรัสสามารถแพร่กระจายผ่านทางช่องระหว่างเซลล์จากเซลล์ไปยังเซลล์ในกรณีนี้จะกลายเป็นน้อยพร้อมที่จะแอนติบอดี
เกณฑ์ทางคลินิกสำหรับการติดเชื้อเอชไอวี
เอชไอวีผู้ใหญ่สร้างถ้าพวกเขามีอย่างน้อยสองอาการร้ายแรงในการรวมกันอย่างน้อยหนึ่งอาการของเล็ก ๆ น้อย ๆ ในกรณีที่ไม่มีสาเหตุอื่น ๆ ที่รู้จักกันของโรคภูมิคุ้มกันบกพร่องที่ (มะเร็ง, โรคภูมิคุ้มกันบกพร่อง แต่กำเนิดความอดอยากอย่างรุนแรงและอื่น ๆ . พี) อาการร้ายแรงรวมถึง:
- การสูญเสียน้ำหนัก 10% หรือมากกว่า;
- ไข้เป็นเวลานานเป็นพัก ๆ หรือหมั่น;
- โรคอุจจาระร่วงเรื้อรัง
อาการเล็กน้อย: ไอถาวร, โรคผิวหนังทั่วไป, งูสวัดที่กำเริบ, candidiasis ของช่องปากและหลอดลม, โรคเริมเรื้อรัง, lymphadenopathy ทั่วไป การวินิจฉัยโรคเอดส์เกิดจากการมีเพียงเยื่อหุ้มสมองของ Kaposi, cryptococcal meningitis, pneumocystis pneumonia ภาพทางคลินิกของโรคนี้ได้รับอิทธิพลจากการติดเชื้อฉวยโอกาส
วิธีการเพาะปลูกไวรัสภูมิคุ้มกันบกพร่องของมนุษย์ (HIV)
HIV-1 และ HIV-2 สามารถเพาะเลี้ยงในเซลล์เดียวของโคลนของ TCB4-lymphocytes - H9 ที่มาจาก leukemic TCV4 lymphocytes วัฒนธรรมแบบแยกตัวของเซลล์ astrocyte สามารถใช้เพื่อวัตถุประสงค์เหล่านี้ซึ่ง HIV-1 คูณกัน จากสัตว์สู่ลิงชิมแปนซีที่ไวต่อการติดเชื้อ HIV-1
ความต้านทานของไวรัสในสิ่งแวดล้อมภายนอกต่ำ เขาเสียชีวิตภายใต้อิทธิพลของแสงแดดและการฉายรังสียูวีถูกทำลายใน 80 องศาเซลเซียสเป็นเวลา 30 นาทีเมื่อใช้สารฆ่าเชื้อที่ใช้กันทั่วไปเป็นเวลา 20-30 นาที ในการฆ่าเชื้อไวรัสที่มีสารจำเป็นต้องใช้สารฆ่าเชื้อ mycobactericidal เนื่องจากมีประสิทธิภาพในการต่อต้านเชื้อจุลินทรีย์ที่มีความต้านทานสูงสุด
การวินิจฉัยโรคในห้องปฏิบัติการของการติดเชื้อเอชไอวี
วิธีหลักในการวินิจฉัยการติดเชื้อไวรัสและเอชไอวีคือเอนไซม์ immunoassay แต่เนื่องจากความจริงที่ว่ามีความคล้ายคลึงกัน gpl20 โครงสร้างและแอนติเจนกับตัวรับของเซลล์ของมนุษย์บางอย่างรวมถึงผู้รับที่ดำเนินการขนส่งของภูมิคุ้มกันบกพร่องผ่านเซลล์เยื่อบุผิวของเยื่อเมือกในร่างกายอาจปรากฏแอนติบอดีแอนติบอดีที่เกี่ยวข้องกับ gpl20 ในกรณีนี้อาจมีผลบวกที่เป็นเท็จของ IFM ดังนั้นทุกซีรั่มที่ทำปฏิกิริยาในเชิงบวกของการศึกษาจะต้องได้รับการวิเคราะห์เพิ่มเติมโดยใช้วิธี immunoblot หรือ western blotting วิธีการนี้ขึ้นอยู่กับการระบุแอนติบอดีที่จะศึกษาหลังจากการแยกสารด้วยไฟฟ้าและการทดสอบในภายหลังโดยใช้แอนติบอดีไวรัสแอนตี้ไวรัส วิธีไวรัสวิทยามีการใช้เพียงเล็กน้อยเนื่องจากความซับซ้อนของการเพาะเชื้อไวรัส โคลนของ H9 lymphocytes ใช้เพื่อให้ได้แอนติเจนไวรัส - ส่วนประกอบที่จำเป็นของระบบการทดสอบเพื่อวินิจฉัย วิธีการ CDR ทำให้สามารถตรวจจับไวรัสได้ในระยะแรกของ viremia
การรักษาผู้ติดเชื้อเอชไอวี
จำเป็นต้องหาหรือสังเคราะห์ยาที่มีประสิทธิภาพในการยับยั้งการทำงานของ reverse transcriptase (revertase) หรือ protease ของไวรัส พวกเขาจะป้องกันการก่อตัวของ DNA provirus และ (หรือ) ยับยั้งการเพิ่มจำนวนของไวรัสภายในเซลล์ กลยุทธ์ที่ทันสมัยของการรักษาผู้ป่วยที่ติดเชื้อ HIV มีพื้นฐานอยู่บนหลักการของการใช้รวมของยาเสพติดที่ยับยั้งเอนไซม์โปรติเอไวรัส (หนึ่งในยาเสพติด) และ revergazu (2 ยาเสพติดที่แตกต่างกัน) ที่ - รวม (สาม) การรักษาด้วย ในรัสเซียสำหรับการรักษาของผู้คนที่ติดเชื้อ HIV ที่แนะนำการใช้งานร่วมกันของสองยาเสพติดในพื้นที่: Crixivan Phosphazide และโดยเฉพาะยับยั้งการสืบพันธุ์ของเอชไอวีในช่วงต้นและปลายของการปรับปรุงพันธุ์โดยเฉพาะอย่างยิ่งกับกิจกรรมที่ลดลงของ AZT
ปัญหาของการป้องกันเฉพาะคือความจำเป็นในการสร้างวัคซีนที่จะช่วยให้เกิดการสร้างภูมิคุ้มกันของเซลล์ที่มีประสิทธิภาพโดยอาศัยเซลล์เม็ดเลือดขาวที่จำเพาะต่อไวรัสโดยไม่มีการผลิตแอนติบอดีที่สำคัญ ภูมิคุ้มกันดังกล่าวจัดทำโดย Thl-helpers เป็นไปได้ว่าแอนติบอดีรวมถึงการ neutralizing ไวรัสไม่เพียง แต่ไม่มีประสิทธิผลในการยับยั้งการติดเชื้อเอชไอวี แต่ในระดับสูงจะช่วยยับยั้งภูมิคุ้มกันในเซลล์ ดังนั้นวัคซีนป้องกันเอชไอวีควรเป็นไปตามข้อกำหนดขั้นพื้นฐานสองประการคือก) ปลอดภัยอย่างเต็มที่และข) กระตุ้นการทำงานของเซลล์เม็ดเลือดขาว T-cytotoxic lymphocytes ศึกษาประสิทธิภาพของตัวแปรต่างๆของวัคซีนที่ได้จากไวรัสที่ตายแล้ว (ที่ไม่ได้ใช้งาน) และจากแอนติเจนแต่ละตัวที่มีคุณสมบัติป้องกันสูง แอนติเจนเหล่านี้สามารถแยกออกจาก virion เองหรือสังเคราะห์ทางเคมีได้ เสนอวัคซีนที่ใช้วิธีการทางพันธุวิศวกรรม เป็นไวรัสฮอร์โมนทดแทนที่มียีน HIV ที่รับผิดชอบในการสังเคราะห์แอนติเจนที่มีคุณสมบัติภูมิคุ้มกันที่แข็งแกร่ง การตัดสินใจเกี่ยวกับประสิทธิผลของวัคซีนเหล่านี้ใช้เวลามากเพราะระยะเวลาการฟักไข่นานในการติดเชื้อเอชไอวีและความแปรปรวนของเชื้อโรคสูง การสร้างวัคซีนป้องกันเอชไอวีที่มีประสิทธิภาพสูงเป็นปัญหาพื้นฐานที่เร่งด่วน