ผู้เชี่ยวชาญทางการแพทย์ของบทความ

สิ่งตีพิมพ์ใหม่
ต่อมใต้สมอง
ตรวจสอบล่าสุด: 04.07.2025

เนื้อหา iLive ทั้งหมดได้รับการตรวจสอบทางการแพทย์หรือตรวจสอบข้อเท็จจริงเพื่อให้แน่ใจว่ามีความถูกต้องตามจริงมากที่สุดเท่าที่จะเป็นไปได้
เรามีแนวทางการจัดหาที่เข้มงวดและมีการเชื่อมโยงไปยังเว็บไซต์สื่อที่มีชื่อเสียงสถาบันการวิจัยทางวิชาการและเมื่อใดก็ตามที่เป็นไปได้ โปรดทราบว่าตัวเลขในวงเล็บ ([1], [2], ฯลฯ ) เป็นลิงก์ที่คลิกได้เพื่อการศึกษาเหล่านี้
หากคุณรู้สึกว่าเนื้อหาใด ๆ ของเราไม่ถูกต้องล้าสมัยหรือมีข้อสงสัยอื่น ๆ โปรดเลือกแล้วกด Ctrl + Enter
ต่อมใต้สมอง (hypophysis, s.glandula pituitaria) ตั้งอยู่ในโพรงใต้สมองของ sella turcica ของกระดูกสฟีนอยด์ และแยกออกจากโพรงกะโหลกศีรษะโดยกระบวนการของดูรามาเตอร์ของสมอง ทำให้เกิดกะบังลมของ sella ต่อมใต้สมองจะเชื่อมต่อกับ infundibulum ของไฮโปทาลามัสของ diencephalon ผ่านช่องเปิดในกะบังลมนี้ ขนาดตามขวางของต่อมใต้สมองคือ 10-17 มม. หน้า-หลังคือ 5-15 มม. แนวตั้งคือ 5-10 มม. มวลของต่อมใต้สมองในผู้ชายคือประมาณ 0.5 กรัม ในผู้หญิงคือ 0.6 กรัม ต่อมใต้สมองถูกปกคลุมภายนอกด้วยแคปซูล
ตามการพัฒนาของต่อมใต้สมองจากรากฐานสองแบบที่แตกต่างกัน กลีบทั้งสองจะถูกแยกออกจากกันในอวัยวะ - กลีบหน้าและกลีบหลัง adenohypophysis หรือกลีบหน้า (adenohypophysis, s.lobus anterior) มีขนาดใหญ่กว่าคิดเป็น 70-80% ของมวลทั้งหมดของต่อมใต้สมอง มีความหนาแน่นมากกว่ากลีบหลัง ในกลีบหน้า มีส่วนปลาย (pars distalis) ซึ่งครอบครองส่วนหน้าของโพรงต่อมใต้สมอง ส่วนกลาง (pars intermedia) ซึ่งอยู่ที่ขอบของกลีบหลัง และส่วนหัว (pars tuberalis) ที่ขึ้นไปและเชื่อมต่อกับ infundibulum ของไฮโปทาลามัส เนื่องจากมีหลอดเลือดมาก กลีบหน้าจึงมีสีเหลืองอ่อนและมีโทนสีแดง เนื้อของต่อมใต้สมองส่วนหน้าประกอบด้วยเซลล์ต่อมหลายประเภท ซึ่งอยู่ระหว่างสายของเซลล์เหล่านี้ซึ่งมีเส้นเลือดฝอยรูปไซนัสซอยด์ตั้งอยู่ เซลล์ต่อมใต้สมองส่วนหน้าครึ่งหนึ่ง (50%) เป็นอะดีโนไซต์ที่ชอบสี ซึ่งมีเม็ดละเอียดในไซโทพลาซึมที่สามารถย้อมด้วยเกลือโครเมียมได้ดี อะดีโนไซต์ที่ชอบสี (40% ของเซลล์ต่อมใต้สมองส่วนหน้าทั้งหมด) และอะดีโนไซต์ที่ชอบสี (10%) อะดีโนไซต์ที่ชอบสี ได้แก่ เอนโดครินไซต์ที่ชอบสี คอร์ติโคโทรปิก และไทรอยด์ อะดีโนไซต์ที่ชอบสีมีขนาดเล็ก มีนิวเคลียสขนาดใหญ่และไซโทพลาซึมในปริมาณเล็กน้อย เซลล์เหล่านี้ถือเป็นสารตั้งต้นของอะดีโนไซต์ที่ชอบสี เซลล์ต่อมใต้สมองส่วนหน้าที่เหลืออีก 50% เป็นอะดีโนไซต์ที่ชอบสี
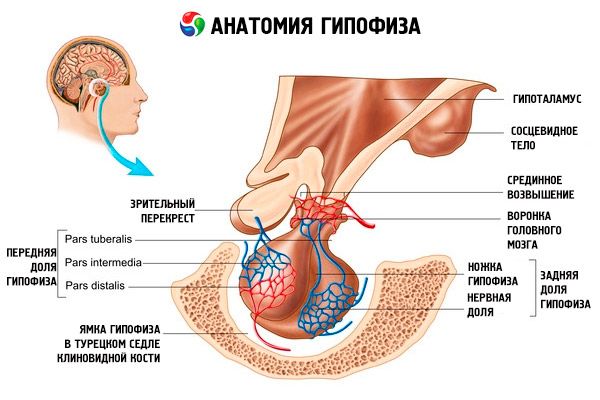
ต่อมใต้สมองส่วนหลัง (neurohypophysis, s.lobus posterior) ประกอบด้วย ต่อมใต้สมองส่วนหลัง (neural lobe) ซึ่งอยู่ที่ส่วนหลังของโพรงต่อมใต้สมอง และกรวย (infundibulum) ซึ่งอยู่ด้านหลังส่วนที่เป็นปุ่มของต่อมใต้สมองส่วนหลัง ต่อมใต้สมองส่วนหลังประกอบด้วยเซลล์ประสาทเกลีย (เซลล์ต่อมใต้สมอง) เส้นใยประสาทที่วิ่งจากนิวเคลียสที่ทำหน้าที่หลั่งสารประสาทของไฮโปทาลามัสไปยังต่อมใต้สมองส่วนหลัง และคอร์พัสเคิลที่ทำหน้าที่หลั่งสารประสาท
ต่อมใต้สมองเชื่อมต่อกับไฮโปทาลามัสของไดเอนเซฟาลอนผ่านทางเส้นประสาท (ทางเดิน) และหลอดเลือด ซึ่งทำหน้าที่ควบคุมการทำงานของต่อมใต้สมอง ต่อมใต้สมองและไฮโปทาลามัสพร้อมกับการเชื่อมต่อระหว่างระบบประสาทต่อมไร้ท่อ หลอดเลือด และเส้นประสาท มักถือเป็นระบบไฮโปทาลามัส-ต่อมใต้สมอง
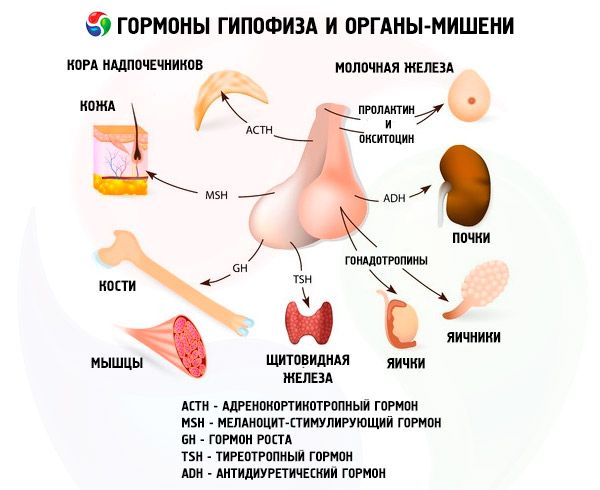
ฮอร์โมนของต่อมใต้สมองส่วนหน้าและส่วนหลังมีผลต่อการทำงานหลายอย่างของร่างกาย โดยส่วนใหญ่ผ่านต่อมไร้ท่ออื่นๆ ในต่อมใต้สมองส่วนหน้า อะดีโนไซต์แอซิโดฟิลิก (เซลล์อัลฟา) จะผลิตฮอร์โมนโซมาโตโทรปิก (ฮอร์โมนการเจริญเติบโต) ซึ่งมีส่วนร่วมในการควบคุมกระบวนการเจริญเติบโตและการพัฒนาของสิ่งมีชีวิตวัยอ่อน เอนโดคริโนไซต์คอร์ติโคโทรปิกจะหลั่งฮอร์โมนอะดรีโนคอร์ติโคโทรปิก (ACTH) ซึ่งกระตุ้นการหลั่งฮอร์โมนสเตียรอยด์ของต่อมหมวกไต เอนโดคริโนไซต์ไทรอยด์โทรปิกจะหลั่งฮอร์โมนไทรอยด์โทรปิก (TSH) ซึ่งส่งผลต่อการพัฒนาของต่อมไทรอยด์และกระตุ้นการผลิตฮอร์โมนของต่อมไทรอยด์ ฮอร์โมนโกนาโดทรอปิก: กระตุ้นการสร้างรูขุมขน (FSH), ลูทีไนซิ่ง (LH) และโพรแลกติน - ส่งผลต่อการเจริญเติบโตทางเพศของร่างกาย ควบคุมและกระตุ้นการพัฒนาของรูขุมขนในรังไข่ การตกไข่ การเจริญเติบโตของต่อมน้ำนมและการผลิตน้ำนมในผู้หญิง กระบวนการสร้างสเปิร์มในผู้ชาย ฮอร์โมนเหล่านี้ผลิตโดยอะดีโนไซต์เบโซฟิลิก (เซลล์เบตา) ปัจจัยไลโปโทรปิกของต่อมใต้สมองยังถูกหลั่งออกมาที่นี่ซึ่งส่งผลต่อการเคลื่อนย้ายและการใช้ไขมันในร่างกาย ในส่วนกลางของกลีบหน้าจะมีการสร้างฮอร์โมนกระตุ้นเมลาโนไซต์ซึ่งควบคุมการสร้างเม็ดสี - เมลานิน - ในร่างกาย
เซลล์ที่หลั่งสารสื่อประสาทของนิวเคลียสเหนือออปติกและพาราเวนทริคิวลาร์ในไฮโปทาลามัสผลิตวาโซเพรสซินและออกซิโทซิน ฮอร์โมนเหล่านี้จะถูกส่งไปยังเซลล์ของต่อมใต้สมองส่วนหลังตามแอกซอนที่ประกอบเป็นทางเดินไฮโปทาลามัส-ต่อมใต้สมอง สารเหล่านี้จะเข้าสู่กระแสเลือดจากต่อมใต้สมองส่วนหลัง ฮอร์โมนวาโซเพรสซินมีฤทธิ์ทำให้หลอดเลือดหดตัวและต่อต้านการขับปัสสาวะ ซึ่งเรียกอีกอย่างว่าฮอร์โมนต่อต้านการขับปัสสาวะ (ADH) ออกซิโทซินมีผลกระตุ้นการหดตัวของกล้ามเนื้อมดลูก เพิ่มการหลั่งน้ำนมของต่อมน้ำนม ยับยั้งการพัฒนาและการทำงานของคอร์ปัสลูเทียม และส่งผลต่อการเปลี่ยนแปลงโทนของกล้ามเนื้อเรียบ (ไม่มีลาย) ของทางเดินอาหาร
การพัฒนาของต่อมใต้สมอง
กลีบหน้าของต่อมใต้สมองพัฒนาจากเยื่อบุผิวของผนังด้านหลังของช่องปากในรูปของส่วนที่ยื่นออกมาเป็นวงแหวน (ถุงของ Rathke) ส่วนที่ยื่นออกมาของชั้นนอกนี้จะเติบโตไปทางด้านล่างของโพรงสมองที่สามในอนาคต กระบวนการจะเติบโตจากพื้นผิวด้านล่างของถุงสมองที่สอง (ซึ่งจะเป็นด้านล่างของโพรงสมองที่สามในอนาคต) ซึ่งตุ่มสีเทาของช่องสมองด้านในและกลีบหลังของต่อมใต้สมองจะพัฒนาจากส่วนนี้
หลอดเลือดและเส้นประสาทของต่อมใต้สมอง
จากหลอดเลือดแดงคาโรติดภายในและหลอดเลือดของวงกลมหลอดเลือดแดงของสมอง หลอดเลือดแดงต่อมใต้สมองส่วนบนและส่วนล่างจะมุ่งไปที่ต่อมใต้สมอง หลอดเลือดแดงต่อมใต้สมองส่วนบนจะมุ่งไปที่นิวเคลียสสีเทาและบริเวณปากเหวของไฮโปทาลามัส โดยเชื่อมต่อกันที่นี่และสร้างเส้นเลือดฝอยที่เจาะเข้าไปในเนื้อเยื่อสมอง ซึ่งก็คือเครือข่ายเส้นเลือดฝอยหลัก จากห่วงยาวและห่วงสั้นของเครือข่ายนี้ จะเกิดเส้นเลือดดำพอร์ทัล ซึ่งมุ่งไปที่กลีบหน้าของต่อมใต้สมอง ในเนื้อของกลีบหน้าของต่อมใต้สมอง เส้นเลือดดำเหล่านี้จะแตกออกเป็นเส้นเลือดฝอยไซนัสกว้าง ก่อให้เกิดเครือข่ายเส้นเลือดฝอยรอง กลีบหลังของต่อมใต้สมองได้รับเลือดจากหลอดเลือดแดงต่อมใต้สมองส่วนล่างเป็นหลัก มีการเชื่อมต่อหลอดเลือดแดงยาวระหว่างหลอดเลือดแดงต่อมใต้สมองส่วนบนและส่วนล่าง เลือดดำที่ไหลออกจากเครือข่ายหลอดเลือดฝอยรองจะเกิดขึ้นผ่านระบบหลอดเลือดดำที่ไหลเข้าสู่โพรงไซนัสและไซนัสระหว่างโพรงไซนัสของเยื่อดูราของสมอง
เส้นใยประสาทซิมพาเทติกที่เข้าสู่อวัยวะพร้อมกับหลอดเลือดแดงมีส่วนร่วมในการส่งสัญญาณของต่อมใต้สมอง เส้นใยประสาทซิมพาเทติกหลังปมประสาทออกจากกลุ่มประสาทของหลอดเลือดแดงคาโรติดภายใน นอกจากนี้ ปลายประสาทจำนวนมากของกระบวนการหลั่งสารประสาทที่อยู่ในนิวเคลียสของไฮโปทาลามัสยังพบในกลีบหลังของต่อมใต้สมองอีกด้วย
ลักษณะต่อมใต้สมองที่เกี่ยวข้องกับอายุ
น้ำหนักเฉลี่ยของต่อมใต้สมองในทารกแรกเกิดอยู่ที่ 0.12 กรัม น้ำหนักของต่อมใต้สมองจะเพิ่มขึ้นเป็นสองเท่าเมื่ออายุ 10 ขวบและเพิ่มขึ้นเป็นสามเท่าเมื่ออายุ 15 ขวบ เมื่ออายุ 20 ขวบ น้ำหนักของต่อมใต้สมองจะถึงจุดสูงสุด (530-560 มก.) และแทบจะไม่เปลี่ยนแปลงในช่วงอายุถัดมา เมื่ออายุ 60 ปี น้ำหนักของต่อมไร้ท่อนี้จะลดลงเล็กน้อย
 [ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
[ 7 ], [ 8 ], [ 9 ], [ 10 ], [ 11 ], [ 12 ], [ 13 ], [ 14 ], [ 15 ]
ฮอร์โมนจากต่อมใต้สมอง
ความสามัคคีของระบบประสาทและการควบคุมฮอร์โมนในร่างกายเกิดขึ้นได้จากการเชื่อมต่อทางกายวิภาคและการทำงานของต่อมใต้สมองและไฮโปทาลามัสอย่างใกล้ชิด คอมเพล็กซ์นี้กำหนดสถานะและการทำงานของระบบต่อมไร้ท่อทั้งหมด
ต่อมไร้ท่อหลักที่ผลิตฮอร์โมนเปปไทด์หลายชนิดซึ่งควบคุมการทำงานของต่อมรอบนอกโดยตรงคือต่อมใต้สมอง ต่อมนี้มีลักษณะเป็นเม็ดสีน้ำตาลแดงคล้ายถั่วมีแคปซูลใยอาหารน้ำหนัก 0.5-0.6 กรัม มีลักษณะแตกต่างกันเล็กน้อยขึ้นอยู่กับเพศและอายุของบุคคล โดยทั่วไปแล้วต่อมใต้สมองแบ่งออกเป็น 2 กลีบซึ่งมีพัฒนาการ โครงสร้าง และหน้าที่ที่แตกต่างกัน ได้แก่ กลีบหน้าส่วนปลาย - ต่อมใต้สมองส่วนหน้าและกลีบหลัง - ต่อมใต้สมองส่วนหลัง กลีบแรกคิดเป็นประมาณ 70% ของมวลรวมของต่อมและโดยทั่วไปจะแบ่งออกเป็นส่วนปลาย กลีบใน และกลีบกลาง ส่วนกลีบหลังหรือกลีบใน และก้านต่อมใต้สมอง ต่อมนี้ตั้งอยู่ในโพรงต่อมใต้สมองของ sella turcica ของกระดูกสฟีนอยด์ และเชื่อมต่อกับสมองผ่านก้าน ส่วนบนของกลีบหน้าถูกปกคลุมด้วยไคแอสมาของเส้นประสาทตาและเส้นประสาทตา เลือดที่ไปเลี้ยงต่อมใต้สมองนั้นมีมากมายและถูกลำเลียงโดยกิ่งก้านของหลอดเลือดแดงคาโรติดภายใน (หลอดเลือดแดงต่อมใต้สมองส่วนบนและส่วนล่าง) เช่นเดียวกับกิ่งก้านของวงหลอดเลือดแดงของสมอง หลอดเลือดแดงต่อมใต้สมองส่วนบนมีส่วนร่วมในการลำเลียงเลือดของต่อมใต้สมองส่วนหน้า และหลอดเลือดแดงที่อยู่ด้านล่าง - ต่อมใต้สมองส่วนหน้า โดยสัมผัสกับปลายของแอกซอนของเซลล์ขนาดใหญ่ในนิวเคลียสของไฮโปทาลามัสที่ทำหน้าที่หลั่งสารสื่อประสาท หลอดเลือดแดงเข้าสู่บริเวณนูนตรงกลางของไฮโปทาลามัส ซึ่งกระจายตัวไปในเครือข่ายหลอดเลือดฝอย (กลุ่มเส้นเลือดฝอยหลัก) เส้นเลือดฝอยเหล่านี้ (ซึ่งเป็นปลายของแอกซอนของเซลล์ประสาทขนาดเล็กที่ทำหน้าที่หลั่งสารสื่อประสาทของไฮโปทาลามัสฐานกลางสัมผัสกับเส้นเลือดฝอย) จะรวมกันเป็นเส้นเลือดดำพอร์ทัลที่ไหลลงตามก้านของต่อมใต้สมองไปยังเนื้อของต่อมใต้สมองส่วนหน้า (adenohypophysis) ซึ่งเส้นเลือดฝอยเหล่านี้จะแยกตัวออกเป็นเครือข่ายของเส้นเลือดฝอยรูปไซนัส (กลุ่มเส้นเลือดฝอยรอง) ดังนั้น เลือดจึงไหลผ่านบริเวณนูนตรงกลางของไฮโปทาลามัส ซึ่งเลือดจะถูกเติมด้วยฮอร์โมนอะดีโนไฮโปฟิโซโทรปิก (ฮอร์โมนปลดปล่อย) ของไฮโปทาลามัส
เลือดที่อิ่มตัวด้วยฮอร์โมนต่อมใต้สมองส่วนหน้าจากเส้นเลือดฝอยจำนวนมากของเส้นประสาทส่วนหน้าจะถูกระบายออกผ่านระบบหลอดเลือดดำ ซึ่งจะไหลเข้าสู่ไซนัสของเยื่อดูราเมเทอร์และเข้าสู่กระแสเลือดทั่วไป ดังนั้น ระบบประตูทางเข้าของต่อมใต้สมองที่มีทิศทางการไหลของเลือดจากไฮโปทาลามัสลงมาจึงเป็นส่วนประกอบทางสัณฐานวิทยาของกลไกที่ซับซ้อนในการควบคุมการทำงานของระบบประสาทและต่อมใต้สมองส่วนหน้า
ต่อมใต้สมองได้รับการควบคุมโดยใยประสาทซิมพาเทติกที่ติดตามหลอดเลือดแดงของต่อมใต้สมอง ใยประสาทเหล่านี้มีต้นกำเนิดจากใยประสาทหลังปมประสาทที่ผ่านกลุ่มเส้นประสาทคาโรติดภายในซึ่งเชื่อมต่อกับปมประสาทส่วนบนของคอ ไม่มีการควบคุมโดยตรงของต่อมใต้สมองส่วนหน้าจากไฮโปทาลามัส กลีบหลังรับใยประสาทจากนิวเคลียสที่หลั่งสารประสาทของไฮโปทาลามัส
ต่อมใต้สมองส่วนหน้าเป็นโครงสร้างทางเนื้อเยื่อที่ซับซ้อนมาก โดยมีเซลล์ต่อมใต้สมอง 2 ประเภท ได้แก่ เซลล์ต่อมใต้สมองที่ไม่ชอบสีและเซลล์ต่อมใต้สมองที่ไม่ชอบสี เซลล์ต่อมใต้สมองส่วนหน้าแบ่งออกเป็นเซลล์ต่อมใต้สมองที่ไม่ชอบสีและเซลล์ต่อมใต้สมองส่วนหน้าชอบสี (คำอธิบายทางเนื้อเยื่อวิทยาโดยละเอียดของต่อมใต้สมองจะระบุไว้ในหัวข้อที่เกี่ยวข้องของคู่มือ) อย่างไรก็ตาม ควรสังเกตว่าฮอร์โมนที่ผลิตโดยเซลล์ต่อมที่ประกอบเป็นเนื้อของต่อมใต้สมองส่วนหน้านั้น มีลักษณะทางเคมีที่แตกต่างกันไปบ้าง เนื่องจากความหลากหลายของเซลล์ต่อมใต้สมองส่วนหน้า และโครงสร้างละเอียดของเซลล์ที่หลั่งฮอร์โมนจะต้องสอดคล้องกับลักษณะการสังเคราะห์ของเซลล์แต่ละเซลล์ แต่บางครั้งเซลล์ต่อมที่สามารถสร้างฮอร์โมนได้หลายชนิดสามารถสังเกตได้ในต่อมใต้สมองส่วนหน้า มีหลักฐานว่าเซลล์ต่อมในต่อมใต้สมองส่วนหน้าไม่ถูกกำหนดทางพันธุกรรมเสมอไป
ใต้กะบังลมของ sella turcica เป็นส่วนที่มีรูปร่างเหมือนกรวยของกลีบหน้า ซึ่งโอบล้อมก้านของต่อมใต้สมองและสัมผัสกับตุ่มสีเทา ส่วนนี้ของต่อมใต้สมองส่วนหน้ามีลักษณะเฉพาะคือมีเซลล์เยื่อบุผิวและมีเลือดไปเลี้ยงมาก นอกจากนี้ยังทำงานด้วยฮอร์โมนอีกด้วย
ส่วนกลางของต่อมใต้สมองประกอบด้วยชั้นเซลล์เบโซฟิลิกขนาดใหญ่ที่ออกฤทธิ์หลั่งสารหลายชั้น
ต่อมใต้สมองทำหน้าที่ต่างๆ ผ่านฮอร์โมน กลีบหน้าผลิตฮอร์โมนอะดรีโนคอร์ติโคโทรปิก (ACTH) ฮอร์โมนกระตุ้นต่อมไทรอยด์ (TSH) ฮอร์โมนกระตุ้นรูขุมขน (FSH) ฮอร์โมนลูทีไนซิง (LH) ฮอร์โมนไลโปโทรปิก รวมถึงฮอร์โมนการเจริญเติบโต เช่น ฮอร์โมนโซมาโทโทรปิก (STO) และฮอร์โมนโพรแลกติน ในกลีบกลาง ฮอร์โมนกระตุ้นเมลาโนไซต์ (MSH) จะถูกสังเคราะห์ และในกลีบหลัง วาสเพรสซินและออกซิโทซินจะสะสม
เอซีทีเอช
ฮอร์โมนต่อมใต้สมองเป็นกลุ่มของโปรตีนและเปปไทด์ฮอร์โมนและไกลโคโปรตีน ในบรรดาฮอร์โมนของต่อมใต้สมองส่วนหน้า ACTH ได้รับการศึกษาวิจัยมาเป็นอย่างดี ฮอร์โมนนี้ผลิตโดยเซลล์เบโซฟิลิก หน้าที่ทางสรีรวิทยาหลักของฮอร์โมนนี้คือการกระตุ้นการสังเคราะห์และการหลั่งฮอร์โมนสเตียรอยด์โดยคอร์เทกซ์ของต่อมหมวกไต ACTH ยังแสดงกิจกรรมกระตุ้นเมลาโนไซต์และไลโปโทรปิก ในปี 1953 ได้มีการแยกออกมาในรูปแบบบริสุทธิ์ ต่อมาได้มีการสร้างโครงสร้างทางเคมีซึ่งประกอบด้วยกรดอะมิโน 39 ชนิดในมนุษย์และสัตว์เลี้ยงลูกด้วยนมหลายชนิด ACTH ไม่มีความจำเพาะต่อสปีชีส์ ปัจจุบันได้มีการสังเคราะห์ฮอร์โมนและชิ้นส่วนต่างๆ ของโมเลกุลทางเคมี ซึ่งมีฤทธิ์มากกว่าฮอร์โมนตามธรรมชาติ โครงสร้างของฮอร์โมนประกอบด้วยห่วงโซ่เปปไทด์สองส่วน ซึ่งส่วนหนึ่งทำหน้าที่ตรวจจับและจับกับตัวรับของ ACTH และอีกส่วนหนึ่งทำหน้าที่ทางชีวภาพ เห็นได้ชัดว่ามันจับกับตัวรับ ACTH ผ่านปฏิกิริยาระหว่างประจุไฟฟ้าของฮอร์โมนและตัวรับ บทบาทของตัวกระตุ้นทางชีวภาพของ ACTH เกิดขึ้นจากชิ้นส่วนของโมเลกุล 4-10 (เมท-กลู-ฮิส-ฟีน-อาร์จี-ไตร-ไตร)
การทำงานของ ACTH ในการกระตุ้นเมลาโนไซต์เกิดจากการมีอยู่ในโมเลกุลของบริเวณปลาย N ซึ่งประกอบด้วยกรดอะมิโน 13 ตัว และมีโครงสร้างซ้ำๆ กันของฮอร์โมนอัลฟาเมลาโนไซต์สติมูเลติ้ง บริเวณนี้ยังมีเฮปตาเปปไทด์ที่มีอยู่ในฮอร์โมนต่อมใต้สมองอื่นๆ และมีกิจกรรมอะดรีโนคอร์ติโคโทรปิก กระตุ้นเมลาโนไซต์ และไลโปโทรปิก
ช่วงเวลาสำคัญในการทำงานของ ACTH ถือเป็นการกระตุ้นเอนไซม์โปรตีนไคเนสในไซโตพลาซึมโดยการมีส่วนร่วมของ cAMP โปรตีนไคเนสที่ถูกฟอสโฟรีเลตจะกระตุ้นเอนไซม์เอสเทอเรส ซึ่งจะเปลี่ยนเอสเทอร์ของคอเลสเตอรอลให้เป็นสารอิสระในหยดไขมัน โปรตีนที่สังเคราะห์ในไซโตพลาซึมอันเป็นผลจากการฟอสโฟรีเลชันของไรโบโซมจะกระตุ้นให้คอเลสเตอรอลอิสระจับกับไซโตโครม P-450 และถ่ายโอนจากหยดไขมันไปยังไมโตคอนเดรีย ซึ่งมีเอนไซม์ทั้งหมดที่รับรองการแปลงคอเลสเตอรอลเป็นคอร์ติโคสเตียรอยด์อยู่
[ 16 ], [ 17 ], [ 18 ], [ 19 ], [ 20 ], [ 21 ], [ 22 ]
ฮอร์โมนกระตุ้นต่อมไทรอยด์
TSH - ไทรอยด์โทรปิน - ตัวควบคุมหลักของการพัฒนาและการทำงานของต่อมไทรอยด์ กระบวนการสังเคราะห์และการหลั่งของฮอร์โมนไทรอยด์ โปรตีนที่ซับซ้อนนี้ - ไกลโคโปรตีน - ประกอบด้วยซับยูนิตอัลฟ่าและเบตา โครงสร้างของซับยูนิตแรกตรงกับซับยูนิตอัลฟ่าของฮอร์โมนลูทีไนซิ่ง นอกจากนี้ ยังตรงกันเป็นส่วนใหญ่ในสัตว์สายพันธุ์ต่างๆ ลำดับของกรดอะมิโนที่ตกค้างในซับยูนิตเบตาของ TSH ของมนุษย์ได้รับการถอดรหัสแล้วและประกอบด้วยกรดอะมิโน 119 ตัว สังเกตได้ว่าซับยูนิตเบตาของ TSH ของมนุษย์และวัวมีความคล้ายคลึงกันในหลายๆ ด้าน คุณสมบัติทางชีวภาพและลักษณะของกิจกรรมทางชีวภาพของฮอร์โมนไกลโคโปรตีนถูกกำหนดโดยซับยูนิตเบตา นอกจากนี้ยังช่วยให้มั่นใจว่าฮอร์โมนมีปฏิสัมพันธ์กับตัวรับในอวัยวะเป้าหมายต่างๆ อย่างไรก็ตาม ซับยูนิตเบตาในสัตว์ส่วนใหญ่แสดงกิจกรรมเฉพาะหลังจากรวมกับซับยูนิตอัลฟ่า ซึ่งทำหน้าที่เป็นตัวกระตุ้นฮอร์โมนชนิดหนึ่ง ปัจจัยหลังนี้มีความน่าจะเป็นเท่ากันในการกระตุ้นให้เกิดกิจกรรม luteinizing, follicle-stimulating และ thyrotropic ซึ่งกำหนดโดยคุณสมบัติของซับยูนิตเบตา ความคล้ายคลึงที่พบช่วยให้เราสรุปได้ว่าฮอร์โมนเหล่านี้เกิดขึ้นในกระบวนการวิวัฒนาการจากบรรพบุรุษร่วมกันตัวหนึ่ง ซับยูนิตเบตายังกำหนดคุณสมบัติทางภูมิคุ้มกันของฮอร์โมนอีกด้วย มีการสันนิษฐานว่าซับยูนิตอัลฟาปกป้องซับยูนิตเบตาจากการทำงานของเอนไซม์โปรตีโอไลติก และยังอำนวยความสะดวกในการขนส่งจากต่อมใต้สมองไปยังอวัยวะเป้าหมายรอบนอกอีกด้วย
ฮอร์โมนโกนาโดโทรปิก
โกนาโดโทรปินมีอยู่ในร่างกายในรูปแบบของ LH และ FSH วัตถุประสงค์ในการทำงานของฮอร์โมนเหล่านี้โดยทั่วไปจะจำกัดอยู่ที่การช่วยในกระบวนการสืบพันธุ์ในบุคคลทั้งสองเพศ พวกมันเป็นโปรตีนที่ซับซ้อน เช่น ไกลโคโปรตีน เช่นเดียวกับ TSH FSH กระตุ้นให้ฟอลลิเคิลในรังไข่ของเพศหญิงเจริญเติบโต และกระตุ้นการสร้างสเปิร์มในเพศชาย LH ทำให้ฟอลลิเคิลแตกในเพศหญิงโดยสร้างคอร์ปัสลูเทียม และกระตุ้นการหลั่งของเอสโตรเจนและโปรเจสเตอโรน ในเพศชาย ฮอร์โมนชนิดเดียวกันจะเร่งการพัฒนาของเนื้อเยื่อระหว่างเซลล์และการหลั่งของแอนโดรเจน ผลของโกนาโดโทรปินขึ้นอยู่กันและเกิดขึ้นพร้อมๆ กัน
พลวัตของการหลั่งฮอร์โมนโกนาโดโทรปินในผู้หญิงจะเปลี่ยนแปลงไปในระหว่างรอบเดือนและได้รับการศึกษาอย่างละเอียดเพียงพอแล้ว ในระยะก่อนการตกไข่ (ฟอลลิเคิล) ของรอบเดือน ปริมาณฮอร์โมน LH จะอยู่ในระดับค่อนข้างต่ำในขณะที่ฮอร์โมน FSH จะเพิ่มขึ้น เมื่อฟอลลิเคิลเจริญเติบโต การหลั่งฮอร์โมนเอสตราไดออลจะเพิ่มขึ้น ส่งผลให้ต่อมใต้สมองผลิตฮอร์โมนโกนาโดโทรปินเพิ่มขึ้น และเกิดรอบเดือนทั้ง LH และ FSH กล่าวคือ สเตียรอยด์เพศจะกระตุ้นการหลั่งฮอร์โมนโกนาโดโทรปิน
ปัจจุบันได้มีการกำหนดโครงสร้างของ LH แล้ว เช่นเดียวกับ TSH ซึ่งประกอบไปด้วยซับยูนิต 2 ตัว คือ a และ b โครงสร้างของซับยูนิตอัลฟาของ LH ในสัตว์แต่ละชนิดนั้นมีความคล้ายคลึงกันเป็นส่วนใหญ่ โดยสอดคล้องกับโครงสร้างของซับยูนิตอัลฟาของ TSH
โครงสร้างของซับยูนิตเบตาของ LH แตกต่างอย่างมากจากโครงสร้างของซับยูนิตเบตาของ TSH แม้ว่าจะมีโซ่เปปไทด์ที่เหมือนกันสี่ส่วนซึ่งประกอบด้วยกรดอะมิโน 4-5 ตัว ใน TSH พวกมันจะอยู่ในตำแหน่ง 27-31, 51-54, 65-68 และ 78-83 เนื่องจากซับยูนิตเบตาของ LH และ TSH กำหนดกิจกรรมทางชีวภาพที่เฉพาะเจาะจงของฮอร์โมน จึงสามารถสันนิษฐานได้ว่าส่วนที่คล้ายคลึงกันในโครงสร้างของ LH และ TSH ควรให้แน่ใจว่าซับยูนิตเบตาเชื่อมโยงกับซับยูนิตอัลฟา และส่วนที่ต่างกันในโครงสร้างควรรับผิดชอบต่อความจำเพาะของกิจกรรมทางชีวภาพของฮอร์โมน
LH ตามธรรมชาติมีเสถียรภาพมากต่อการทำงานของเอนไซม์โปรตีโอไลติก แต่ซับยูนิตเบตาจะถูกตัดออกอย่างรวดเร็วโดยไคโมทริปซิน และซับยูนิต a นั้นยากต่อการไฮโดรไลซ์โดยเอนไซม์ กล่าวคือ มีบทบาทในการปกป้องโดยป้องกันไม่ให้ไคโมทริปซินเข้าถึงพันธะเปปไทด์
นักวิจัยยังไม่สามารถสรุปโครงสร้างทางเคมีของ FSH ได้อย่างชัดเจน เช่นเดียวกับ LH FSH ประกอบด้วยซับยูนิต 2 ยูนิต แต่ซับยูนิตเบตาของ FSH แตกต่างจากซับยูนิตเบตาของ LH
โพรแลกติน
ฮอร์โมนอีกชนิดหนึ่งคือ โพรแลกติน (ฮอร์โมนแลคโตเจน) มีบทบาทสำคัญในกระบวนการสืบพันธุ์ คุณสมบัติทางสรีรวิทยาหลักของโพรแลกตินในสัตว์เลี้ยงลูกด้วยนมนั้นแสดงออกมาในรูปแบบของการกระตุ้นการพัฒนาของต่อมน้ำนมและการให้นม การเจริญเติบโตของต่อมไขมันและอวัยวะภายใน โพรแลกตินส่งเสริมการแสดงออกของผลของสเตียรอยด์ต่อลักษณะทางเพศรองในตัวผู้ กระตุ้นกิจกรรมการหลั่งของคอร์ปัสลูเทียมในหนูและหนูตะเภา และมีส่วนร่วมในการควบคุมการเผาผลาญไขมัน ในช่วงไม่กี่ปีที่ผ่านมา โพรแลกตินได้รับความสนใจอย่างมากในฐานะตัวควบคุมพฤติกรรมการเป็นแม่ การทำงานหลายอย่างดังกล่าวอธิบายได้จากการพัฒนาเชิงวิวัฒนาการ โพรแลกตินเป็นฮอร์โมนต่อมใต้สมองโบราณชนิดหนึ่งและพบได้แม้แต่ในสัตว์สะเทินน้ำสะเทินบก ปัจจุบัน โครงสร้างของโพรแลกตินในสัตว์เลี้ยงลูกด้วยนมบางสายพันธุ์ได้รับการถอดรหัสอย่างสมบูรณ์แล้ว อย่างไรก็ตาม จนกระทั่งเมื่อไม่นานนี้ นักวิทยาศาสตร์ยังคงแสดงความสงสัยเกี่ยวกับการมีอยู่ของฮอร์โมนดังกล่าวในมนุษย์ หลายคนเชื่อว่าหน้าที่ของโพรแลกตินนั้นเกิดจากฮอร์โมนการเจริญเติบโต ปัจจุบันมีหลักฐานที่น่าเชื่อถือเกี่ยวกับการปรากฏตัวของโพรแลกตินในมนุษย์ และโครงสร้างของโพรแลกตินได้รับการถอดรหัสบางส่วนแล้ว ตัวรับโพรแลกตินจับกับฮอร์โมนการเจริญเติบโตและแล็กโตเจนของรกอย่างแข็งขัน ซึ่งบ่งชี้ถึงกลไกการทำงานของฮอร์โมนทั้งสามชนิด
โซมาโทโทรปิน
ฮอร์โมนการเจริญเติบโตหรือโซมาโทโทรปินมีขอบเขตการออกฤทธิ์ที่กว้างกว่าโพรแลกติน เช่นเดียวกับโพรแลกติน ฮอร์โมนนี้ถูกผลิตโดยเซลล์แอซิโดฟิลิกของต่อมใต้สมองส่วนหน้า (adenohypophysis) ฮอร์โมนการเจริญเติบโตจะกระตุ้นการเจริญเติบโตของโครงกระดูก กระตุ้นการสังเคราะห์โปรตีน มีผลในการเคลื่อนย้ายไขมัน และส่งเสริมให้ขนาดร่างกายเพิ่มขึ้น นอกจากนี้ยังช่วยประสานงานกระบวนการเผาผลาญอีกด้วย
การมีส่วนร่วมของฮอร์โมนในช่วงหลังนี้ได้รับการยืนยันจากข้อเท็จจริงของการหลั่งที่เพิ่มขึ้นอย่างรวดเร็วโดยต่อมใต้สมอง เช่น เมื่อระดับน้ำตาลในเลือดลดลง
โครงสร้างทางเคมีของฮอร์โมนของมนุษย์นี้ได้รับการสร้างเสร็จสมบูรณ์แล้ว โดยมีกรดอะมิโน 191 ตัว โครงสร้างหลักของฮอร์โมนนี้คล้ายคลึงกับโครงสร้างของฮอร์โมนโคริโอนิกโซมาโตแมมโมโทรปินหรือแล็กโตเจนของรก ข้อมูลเหล่านี้บ่งชี้ถึงความใกล้ชิดทางวิวัฒนาการที่สำคัญของฮอร์โมนทั้งสองชนิด แม้ว่าจะมีความแตกต่างกันในกิจกรรมทางชีวภาพก็ตาม
จำเป็นต้องเน้นย้ำถึงความจำเพาะสูงของสายพันธุ์ของฮอร์โมนที่เป็นปัญหา - ตัวอย่างเช่น STH ที่มีต้นกำเนิดจากสัตว์ไม่ทำงานในมนุษย์ สิ่งนี้ได้รับการอธิบายโดยทั้งปฏิกิริยาระหว่างตัวรับของ STH ของมนุษย์และสัตว์และโครงสร้างของฮอร์โมนเอง ปัจจุบันกำลังศึกษาวิจัยเพื่อระบุศูนย์ที่ทำงานในโครงสร้างที่ซับซ้อนของ STH ที่แสดงกิจกรรมทางชีวภาพ ชิ้นส่วนแต่ละชิ้นของโมเลกุลที่แสดงคุณสมบัติอื่นๆ ได้รับการศึกษา ตัวอย่างเช่น หลังจากการไฮโดรไลซิส STH ของมนุษย์โดยเปปซิน เปปไทด์ที่ประกอบด้วยกรดอะมิโน 14 ตัวและสอดคล้องกับส่วนโมเลกุล 31-44 ได้ถูกแยกออก เปปไทด์นี้ไม่มีผลต่อการเจริญเติบโต แต่มีประสิทธิภาพมากกว่าฮอร์โมนดั้งเดิมอย่างมีนัยสำคัญในกิจกรรมไลโปโทรปิก ฮอร์โมนการเจริญเติบโตของมนุษย์ซึ่งแตกต่างจากฮอร์โมนที่คล้ายกันของสัตว์มีกิจกรรมแลคโตเจนิกอย่างมีนัยสำคัญ
ต่อมใต้สมองส่วนหน้าสังเคราะห์สารเปปไทด์และโปรตีนหลายชนิดที่มีผลในการเคลื่อนย้ายไขมัน และฮอร์โมนต่อมใต้สมองส่วนหน้า - ACTH, STH, TSH และอื่นๆ - มีผลไลโปโทรปิก ในช่วงไม่กี่ปีที่ผ่านมา ฮอร์โมนเบตาและวาย-ไลโปโทรปิก (LPG) ได้รับการเน้นย้ำเป็นพิเศษ คุณสมบัติทางชีวภาพของเบตา-LPG ได้รับการศึกษาอย่างละเอียดถี่ถ้วน นอกจากกิจกรรมไลโปโทรปิกแล้ว ยังมีผลกระตุ้นเมลาโนไซต์ กระตุ้นคอร์ติโคโทรปิน และไฮโปแคลซีเมีย และยังก่อให้เกิดผลคล้ายอินซูลินอีกด้วย
ปัจจุบันโครงสร้างหลักของ LPG ในแกะ (กรดอะมิโน 90 ตัว) ฮอร์โมนไลโปโทรปิกของหมูและวัวได้รับการถอดรหัสแล้ว ฮอร์โมนนี้มีความเฉพาะเจาะจงต่อสายพันธุ์ แม้ว่าโครงสร้างของบริเวณกลางของเบตา-LPG จะเหมือนกันในสายพันธุ์ต่าง ๆ ก็ตาม ฮอร์โมนนี้กำหนดคุณสมบัติทางชีวภาพของฮอร์โมน หนึ่งในชิ้นส่วนของบริเวณนี้พบได้ในโครงสร้างของอัลฟา-MSH เบตา-MSH ACTH และเบตา-LPG มีการเสนอว่าฮอร์โมนเหล่านี้เกิดจากสารตั้งต้นเดียวกันในกระบวนการวิวัฒนาการ γ-LPG มีกิจกรรมไลโปโทรปิกที่อ่อนแอกว่าเบตา-LPG
ฮอร์โมนกระตุ้นเมลาโนไซต์
ฮอร์โมนนี้สังเคราะห์ในกลีบกลางของต่อมใต้สมอง กระตุ้นการสังเคราะห์เมลานินซึ่งเป็นเม็ดสีของผิวหนังในการทำงานทางชีวภาพ ส่งเสริมให้ขนาดและจำนวนของเซลล์เม็ดสีเมลาโนไซต์ในผิวหนังของสัตว์สะเทินน้ำสะเทินบกเพิ่มขึ้น คุณสมบัติเหล่านี้ของ MSH ใช้ในการทดสอบทางชีววิทยาของฮอร์โมน ฮอร์โมนมีสองประเภทคือ อัลฟาและเบตา-MSH ได้รับการพิสูจน์แล้วว่าอัลฟา-MSH ไม่มีความจำเพาะต่อสายพันธุ์และมีโครงสร้างทางเคมีเหมือนกันในสัตว์เลี้ยงลูกด้วยนมทั้งหมด โมเลกุลของมันคือโซ่เปปไทด์ที่ประกอบด้วยกรดอะมิโน 13 ตัว ในทางกลับกัน เบตา-MSH มีความจำเพาะต่อสายพันธุ์ และโครงสร้างของมันแตกต่างกันไปในสัตว์แต่ละชนิด ในสัตว์เลี้ยงลูกด้วยนมส่วนใหญ่ โมเลกุลของเบตา-MSH ประกอบด้วยกรดอะมิโน 18 ตัว และในมนุษย์เท่านั้นที่ขยายออกจากปลายกรดอะมิโนด้วยกรดอะมิโน 4 ตัว ควรสังเกตว่าอัลฟา-MSH มีกิจกรรมอะดรีโนคอร์ติโคโทรปิก และผลกระทบต่อพฤติกรรมของสัตว์และมนุษย์ได้รับการพิสูจน์แล้วในปัจจุบัน
ออกซิโทซินและวาสเพรสซิน
วาโซเพรสซินและออกซิโทซินซึ่งสังเคราะห์ในไฮโปทาลามัสจะสะสมในกลีบหลังของต่อมใต้สมอง โดยวาโซเพรสซินจะอยู่ในนิวรอนของนิวเคลียสซูพราออปติก และออกซิโทซินในนิวเคลียสพาราเวนตริคูเลอรี่ จากนั้นสารเหล่านี้จะถูกส่งไปยังต่อมใต้สมอง ควรเน้นว่าสารตั้งต้นของฮอร์โมนวาโซเพรสซินจะถูกสังเคราะห์ในไฮโปทาลามัสก่อน ในเวลาเดียวกัน โปรตีนนิวโรฟิซินชนิด 1 และ 2 จะถูกผลิตที่นั่น โปรตีนชนิดแรกจะจับกับออกซิโทซิน และชนิดที่สองจะจับกับวาโซเพรสซิน คอมเพล็กซ์เหล่านี้จะอพยพในรูปแบบของแกรนูลที่หลั่งสารประสาทในไซโทพลาซึมตามแนวแอกซอนและไปถึงกลีบหลังของต่อมใต้สมอง ซึ่งเส้นประสาทจะสิ้นสุดที่ผนังหลอดเลือด และเนื้อหาของแกรนูลจะเข้าสู่กระแสเลือด วาโซเพรสซินและออกซิโทซินเป็นฮอร์โมนต่อมใต้สมองตัวแรกที่มีลำดับกรดอะมิโนที่ถูกกำหนดไว้อย่างสมบูรณ์ ในโครงสร้างทางเคมี ฮอร์โมนทั้งสองเป็นโนนาเปปไทด์ที่มีสะพานไดซัลไฟด์หนึ่งแห่ง
ฮอร์โมนที่อยู่ระหว่างการพิจารณามีผลทางชีวภาพต่างๆ เช่น กระตุ้นการขนส่งน้ำและเกลือผ่านเยื่อหุ้มเซลล์ มีฤทธิ์กระตุ้นหลอดเลือด เพิ่มการหดตัวของกล้ามเนื้อเรียบของมดลูกระหว่างการคลอดบุตร และเพิ่มการหลั่งของต่อมน้ำนม ควรสังเกตว่าวาโซเพรสซินมีฤทธิ์ต้านการขับปัสสาวะมากกว่าออกซิโทซิน ในขณะที่ออกซิโทซินมีผลต่อมดลูกและต่อมน้ำนมมากกว่า ตัวควบคุมการหลั่งวาโซเพรสซินหลักคือการบริโภคน้ำ ในหลอดไต วาโซเพรสซินจะจับกับตัวรับในเยื่อหุ้มไซโทพลาสซึม จากนั้นจึงกระตุ้นเอนไซม์อะดีไนเลตไซเคลสในเยื่อหุ้มไซโทพลาสซึม ส่วนต่างๆ ของโมเลกุลมีหน้าที่จับฮอร์โมนกับตัวรับและมีผลทางชีวภาพ
ต่อมใต้สมองซึ่งเชื่อมต่อผ่านไฮโปทาลามัสกับระบบประสาททั้งหมดจะรวมระบบต่อมไร้ท่อให้เป็นหนึ่งเดียวที่ทำงานได้ โดยมีส่วนร่วมในการทำให้สภาพแวดล้อมภายในร่างกายมีความคงที่ (โฮมีโอสตาซิส) ภายในระบบต่อมไร้ท่อ การควบคุมโฮมีโอสตาซิสจะดำเนินการตามหลักการของการตอบรับระหว่างกลีบหน้าของต่อมใต้สมองและต่อม "เป้าหมาย" (ต่อมไทรอยด์ คอร์เทกซ์ต่อมหมวกไต ต่อมเพศ) ฮอร์โมนส่วนเกินที่ผลิตโดยต่อม "เป้าหมาย" จะยับยั้ง และการขาดฮอร์โมนจะกระตุ้นการหลั่งและการปลดปล่อยฮอร์โมนโทรปิกที่เกี่ยวข้อง ไฮโปทาลามัสรวมอยู่ในระบบตอบรับ ซึ่งเป็นที่ตั้งโซนตัวรับที่ไวต่อฮอร์โมนของต่อม "เป้าหมาย" โดยการจับกับฮอร์โมนที่ไหลเวียนอยู่ในเลือดโดยเฉพาะและเปลี่ยนแปลงการตอบสนองขึ้นอยู่กับความเข้มข้นของฮอร์โมน ตัวรับไฮโปทาลามัสจะถ่ายทอดผลไปยังศูนย์ไฮโปทาลามัสที่เกี่ยวข้อง ซึ่งประสานงานการทำงานของต่อมใต้สมองส่วนหน้าและต่อมใต้สมองส่วนหน้า โดยปล่อยฮอร์โมนต่อมใต้สมองส่วนหน้าและต่อมใต้สมองส่วนหน้า ดังนั้น จึงควรพิจารณาว่าไฮโปทาลามัสเป็นสมองต่อมไร้ท่อประสาท