ผู้เชี่ยวชาญทางการแพทย์ของบทความ

สิ่งตีพิมพ์ใหม่
การสังเคราะห์ การหลั่ง และการเผาผลาญของคาเทโคลามีน
ตรวจสอบล่าสุด: 06.07.2025

เนื้อหา iLive ทั้งหมดได้รับการตรวจสอบทางการแพทย์หรือตรวจสอบข้อเท็จจริงเพื่อให้แน่ใจว่ามีความถูกต้องตามจริงมากที่สุดเท่าที่จะเป็นไปได้
เรามีแนวทางการจัดหาที่เข้มงวดและมีการเชื่อมโยงไปยังเว็บไซต์สื่อที่มีชื่อเสียงสถาบันการวิจัยทางวิชาการและเมื่อใดก็ตามที่เป็นไปได้ โปรดทราบว่าตัวเลขในวงเล็บ ([1], [2], ฯลฯ ) เป็นลิงก์ที่คลิกได้เพื่อการศึกษาเหล่านี้
หากคุณรู้สึกว่าเนื้อหาใด ๆ ของเราไม่ถูกต้องล้าสมัยหรือมีข้อสงสัยอื่น ๆ โปรดเลือกแล้วกด Ctrl + Enter
ต่อมหมวกไตส่วนในสร้างสารประกอบที่มีโครงสร้างที่แตกต่างจากสเตียรอยด์ สารประกอบเหล่านี้ประกอบด้วยนิวเคลียส 3,4-dihydroxyphenyl (catechol) และเรียกว่า catecholamine ซึ่งได้แก่ อะดรีนาลีน นอร์เอพิเนฟริน และโดพามีน เบตาไฮดรอกซีไทรามีน
ลำดับการสังเคราะห์ catecholamine นั้นค่อนข้างง่าย: ไทโรซีน → ไดไฮดรอกซีฟีนิลอะลานีน (DOPA) → โดปามีน → นอร์เอพิเนฟริน → อะดรีนาลีน ไทโรซีนเข้าสู่ร่างกายพร้อมกับอาหาร แต่สามารถสร้างได้จากฟีนิลอะลานีนในตับภายใต้การทำงานของฟีนิลอะลานีนไฮดรอกซิเลส ผลิตภัณฑ์ขั้นสุดท้ายของการแปลงไทโรซีนในเนื้อเยื่อนั้นแตกต่างกัน ในเมดัลลาของต่อมหมวกไต กระบวนการดำเนินไปจนถึงขั้นตอนการสร้างอะดรีนาลีน ที่ปลายประสาทซิมพาเทติก - นอร์เอพิเนฟริน ในเซลล์ประสาทบางส่วนของระบบประสาทส่วนกลาง การสังเคราะห์ catecholamine จะสิ้นสุดลงด้วยการสร้างโดปามีน
การเปลี่ยนไทโรซีนเป็นโดปาเกิดขึ้นโดยไทโรซีนไฮดรอกซิเลส ซึ่งมีโคแฟกเตอร์คือเทตระไฮโดรไบโอพเทอรินและออกซิเจน เชื่อกันว่าเอนไซม์นี้จำกัดอัตราของกระบวนการทั้งหมดของการสังเคราะห์คาเทโคลามีน และถูกยับยั้งโดยผลิตภัณฑ์ขั้นสุดท้ายของกระบวนการ ไทโรซีนไฮดรอกซิเลสเป็นเป้าหมายหลักของผลควบคุมต่อการสังเคราะห์คาเทโคลามีน
การเปลี่ยน DOPA เป็นโดพามีนเกิดขึ้นจากเอนไซม์ DOPA decarboxylase (โคแฟกเตอร์ - ไพริดอกซัลฟอสเฟต) ซึ่งค่อนข้างไม่จำเพาะและดีคาร์บอกซิเลตกรดอะมิโน L อะโรมาติกอื่นๆ อย่างไรก็ตาม มีข้อบ่งชี้ถึงความเป็นไปได้ในการปรับเปลี่ยนการสังเคราะห์คาเทโคลามีนโดยการเปลี่ยนกิจกรรมของเอนไซม์นี้ เซลล์ประสาทบางส่วนขาดเอนไซม์สำหรับการแปลงโดพามีนเพิ่มเติม และเป็นผลิตภัณฑ์ขั้นสุดท้าย เนื้อเยื่ออื่นๆ มีโดพามีนเบตาไฮดรอกซีเลส (โคแฟกเตอร์ - ทองแดง กรดแอสคอร์บิก และออกซิเจน) ซึ่งแปลงโดพามีนเป็นนอร์เอพิเนฟริน ในเมดัลลาของต่อมหมวกไต (แต่ไม่ใช่ที่ปลายของเส้นประสาทซิมพาเทติก) มีฟีนิลเอทาโนลามีนอยู่ ซึ่งเป็นเมทิลทรานสเฟอเรสที่สร้างอะดรีนาลีนจากนอร์เอพิเนฟริน ในกรณีนี้ S-adenosylmethionine ทำหน้าที่เป็นผู้บริจาคกลุ่มเมทิล
สิ่งสำคัญที่ต้องจำไว้คือ การสังเคราะห์ฟีนิลเอทาโนลามีน-เอ็น-เมทิลทรานสเฟอเรสถูกเหนี่ยวนำโดยกลูโคคอร์ติคอยด์ที่เข้าสู่เมดัลลาจากคอร์เทกซ์ผ่านระบบหลอดเลือดดำพอร์ทัล ซึ่งอาจอธิบายได้ว่าทำไมต่อมไร้ท่อสองต่อมที่ต่างกันจึงรวมกันอยู่ในอวัยวะเดียวกัน ความสำคัญของกลูโคคอร์ติคอยด์สำหรับการสังเคราะห์อะดรีนาลีนนั้นเน้นย้ำจากข้อเท็จจริงที่ว่าเซลล์ของเมดัลลาต่อมหมวกไตที่ผลิตนอร์เอพิเนฟรินนั้นอยู่รอบ ๆ หลอดเลือดแดง ในขณะที่เซลล์ที่ผลิตอะดรีนาลีนจะได้รับเลือดส่วนใหญ่จากไซนัสหลอดเลือดดำซึ่งอยู่ในคอร์เทกซ์ต่อมหมวกไต
การสลายตัวของคาเทโคลามีนเกิดขึ้นส่วนใหญ่ภายใต้อิทธิพลของระบบเอนไซม์สองระบบ ได้แก่ คาเทโคลามีน-โอ-เมทิลทรานสเฟอเรส (COMT) และโมโนเอมีนออกซิเดส (MAO) เส้นทางหลักของการสลายตัวของอะดรีนาลีนและนอร์เอพิเนฟรินแสดงไว้โดยแผนผังในรูปที่ 54 ภายใต้อิทธิพลของ COMT ที่มีตัวให้กลุ่มเมทิล S-adrenosylmethionine คาเทโคลามีนจะถูกแปลงเป็นนอร์เมทาเนฟรินและเมทาเนฟริน (อนุพันธ์ 3-O-เมทิลของนอร์เอพิเนฟรินและอะดรีนาลีน) ซึ่งภายใต้อิทธิพลของ MAO จะถูกแปลงเป็นอัลดีไฮด์และจากนั้น (ภายใต้ตัวให้อัลดีไฮด์ออกซิเดส) เป็นกรดวานิลลิลแมนเดลิก (VMA) ซึ่งเป็นผลิตภัณฑ์การสลายหลักของนอร์เอพิเนฟรินและอะดรีนาลีน ในกรณีเดียวกัน เมื่อคาเทโคลามีนสัมผัสกับ MAO ในตอนแรกแทนที่จะเป็น COMT คาเทโคลามีนจะถูกแปลงเป็น 3,4-dioxomandelic aldehyde จากนั้นภายใต้อิทธิพลของ aldehyde oxidase และ COMT จะกลายเป็น 3,4-dioxomandelic acid และ VMC ในสภาวะที่มีแอลกอฮอล์ดีไฮโดรจีเนส 3-methoxy-4-oxyphenylglycol ซึ่งเป็นผลิตภัณฑ์ขั้นสุดท้ายหลักของการย่อยสลายอะดรีนาลีนและนอร์เอพิเนฟรินใน CNS สามารถสร้างขึ้นได้จากคาเทโคลามีน
การสลายตัวของโดปามีนมีความคล้ายคลึงกัน ยกเว้นว่าเมแทบอไลต์ของมันขาดกลุ่มไฮดรอกซิลที่อะตอมเบตาคาร์บอน ดังนั้นจึงเกิดกรดโฮโมวานิลลิก (HVA) หรือกรด 3-เมทอกซี-4-ไฮดรอกซีฟีนิลอะซิติกแทนที่จะเป็นกรดวานิลลิลแมนเดลิก
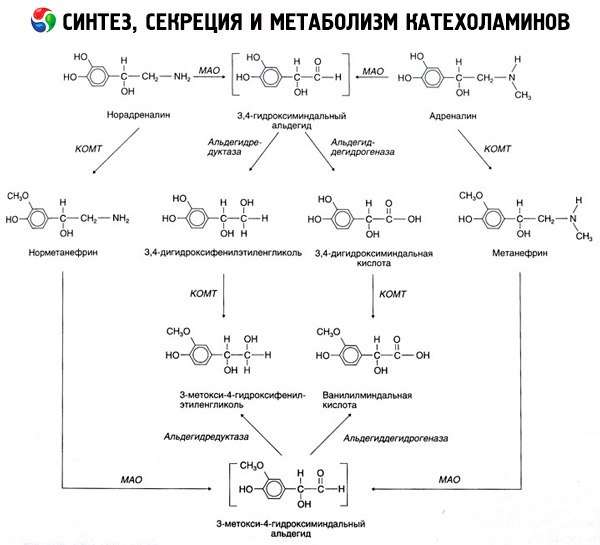
ยังมีการตั้งสมมติฐานเกี่ยวกับการดำรงอยู่ของเส้นทางควิโนอิดของออกซิเดชันของโมเลกุลคาเทโคลามีน ซึ่งอาจผลิตผลิตภัณฑ์กลางที่มีกิจกรรมทางชีวภาพที่เด่นชัด
นอร์เอพิเนฟรินและอะดรีนาลีนที่ก่อตัวขึ้นภายใต้การทำงานของเอนไซม์ไซโทซอลในปลายประสาทซิมพาเทติกและต่อมหมวกไตส่วนในจะเข้าสู่แกรนูลหลั่งสารซึ่งจะปกป้องแกรนูลเหล่านี้จากการทำงานของเอนไซม์ย่อยสลาย การจับคาเทโคลามีนโดยแกรนูลต้องใช้พลังงาน ในแกรนูลโครมาฟฟินของต่อมหมวกไตส่วนใน คาเทโคลามีนจะจับกับ ATP อย่างแน่นหนา (ในอัตราส่วน 4:1) และโปรตีนเฉพาะ - โครโมแกรนิน ซึ่งป้องกันการแพร่กระจายของฮอร์โมนจากแกรนูลเข้าไปในไซโทพลาสซึม
การกระตุ้นโดยตรงสำหรับการหลั่ง catecholamine ดูเหมือนว่าจะเป็นการแทรกซึมของแคลเซียมเข้าไปในเซลล์ กระตุ้นการขับออกนอกเซลล์ (การหลอมรวมของเยื่อเม็ดกับพื้นผิวเซลล์และการแตกออกพร้อมกับการปล่อยเนื้อหาที่ละลายได้ทั้งหมด - catecholamine, โดพามีนเบตาไฮดรอกซิเลส, ATP และโครโมแกรนิน - ลงในของเหลวนอกเซลล์)
ผลทางสรีรวิทยาของคาเทโคลามีนและกลไกการออกฤทธิ์
ผลกระทบของคาเทโคลามีนเริ่มต้นจากการโต้ตอบกับตัวรับเฉพาะของเซลล์เป้าหมาย ในขณะที่ตัวรับฮอร์โมนไทรอยด์และสเตียรอยด์อยู่ภายในเซลล์ ตัวรับคาเทโคลามีน (เช่นเดียวกับฮอร์โมนอะเซทิลโคลีนและเปปไทด์) จะอยู่บนพื้นผิวเซลล์ด้านนอก
ได้มีการพิสูจน์มานานแล้วว่าอะดรีนาลีนหรือนอร์เอพิเนฟรินมีประสิทธิภาพมากกว่าคาเทโคลามีนไอโซโพรเทอเรนอลสังเคราะห์ในปฏิกิริยาบางอย่าง ในขณะที่ไอโซโพรเทอเรนอลมีประสิทธิภาพเหนือกว่าอะดรีนาลีนหรือนอร์เอพิเนฟรินในปฏิกิริยาอื่นๆ จากพื้นฐานนี้ แนวคิดเกี่ยวกับการมีอยู่ของอะดรีนาลีนรีเซพเตอร์สองประเภทในเนื้อเยื่อ ได้แก่ อัลฟาและเบตา และในเนื้อเยื่อบางส่วนจะมีได้เพียงประเภทเดียวเท่านั้น ไอโซโพรเทอเรนอลเป็นอะโกนิสต์ที่ทรงพลังที่สุดของเบตา-อะดรีนาลีนรีเซพเตอร์ ในขณะที่ฟีนิลเอฟรินซึ่งเป็นสารประกอบสังเคราะห์เป็นอะโกนิสต์ที่ทรงพลังที่สุดของอัลฟา-อะดรีนาลีน คาเทโคลามีนจากธรรมชาติ ได้แก่ อะดรีนาลีนและนอร์เอพิเนฟริน สามารถโต้ตอบกับตัวรับทั้งสองประเภทได้ แต่อะดรีนาลีนมีความสัมพันธ์กับเบตาและนอร์เอพิเนฟรินมากกว่า
คาเทโคลามีนกระตุ้นตัวรับเบต้า-อะดรีเนอร์จิกของหัวใจได้แรงกว่าตัวรับเบต้า-อะดรีเนอร์จิกของกล้ามเนื้อเรียบ ซึ่งทำให้สามารถแบ่งตัวรับเบต้าออกเป็นประเภทย่อยได้ ได้แก่ ตัวรับเบต้า 1 (หัวใจ เซลล์ไขมัน) และตัวรับเบต้า 2 (หลอดลม หลอดเลือด ฯลฯ) ผลของไอโซโพรเทอเรนอลต่อตัวรับเบต้า 1 นั้นมากกว่าผลของอะดรีนาลีนและนอร์เอพิเนฟรินเพียง 10 เท่า ในขณะที่ไอโซโพรเทอเรนอลออกฤทธิ์แรงกว่าคาเทโคลามีนจากธรรมชาติถึง 100-1,000 เท่าต่อตัวรับเบต้า 2
การใช้สารต่อต้านเฉพาะ (phentolamine และ phenoxybenzamine สำหรับอัลฟาและ propranolol สำหรับเบตารีเซพเตอร์) ยืนยันความเหมาะสมของการจำแนกประเภทของตัวรับอะดรีโน โดพามีนสามารถโต้ตอบกับตัวรับอัลฟาและเบตาได้ แต่เนื้อเยื่อต่างๆ (สมอง ต่อมใต้สมอง หลอดเลือด) ก็มีตัวรับโดพามีนของตัวเองเช่นกัน ซึ่งตัวบล็อกเฉพาะคือฮาโลเพอริดอล จำนวนเบตารีเซพเตอร์จะแตกต่างกันไปตั้งแต่ 1,000 ถึง 2,000 ตัวต่อเซลล์ ผลทางชีวภาพของคาเทโคลามีนที่ควบคุมโดยเบตารีเซพเตอร์มักเกี่ยวข้องกับการกระตุ้นอะดีไนเลตไซเคลสและการเพิ่มขึ้นของเนื้อหาภายในเซลล์ของ cAMP แม้ว่าตัวรับและเอนไซม์จะเชื่อมต่อกันทางหน้าที่ แต่ก็เป็นโมเลกุลขนาดใหญ่ที่แตกต่างกัน กัวโนซีนไตรฟอสเฟต (GTP) และนิวคลีโอไทด์พิวรีนอื่นๆ มีส่วนร่วมในการปรับกิจกรรมอะดีไนเลตไซเคลสภายใต้อิทธิพลของคอมเพล็กซ์ฮอร์โมนรีเซพเตอร์ การเพิ่มกิจกรรมของเอนไซม์ทำให้ความสัมพันธ์ของตัวรับเบตากับสารก่อฤทธิ์ลดลง
ปรากฏการณ์ของความไวที่เพิ่มขึ้นของโครงสร้างที่ถูกตัดเส้นประสาทเป็นที่ทราบกันมานานแล้ว ในทางตรงกันข้าม การสัมผัสกับสารกระตุ้นเป็นเวลานานจะลดความไวของเนื้อเยื่อเป้าหมาย การศึกษาเกี่ยวกับตัวรับเบตาทำให้สามารถอธิบายปรากฏการณ์เหล่านี้ได้ มีการแสดงให้เห็นว่าการสัมผัสกับไอโซโพรเทอเรนอลเป็นเวลานานนำไปสู่การสูญเสียความไวของอะดีไนเลตไซเคลสเนื่องจากจำนวนตัวรับเบตาลดลง
กระบวนการลดความไวต่อสิ่งเร้าไม่จำเป็นต้องมีการกระตุ้นการสังเคราะห์โปรตีน และอาจเกิดจากการสร้างคอมเพล็กซ์ตัวรับฮอร์โมนที่ไม่สามารถย้อนกลับได้ขึ้นอย่างค่อยเป็นค่อยไป ในทางกลับกัน การนำ 6-oxidopamine เข้ามา ซึ่งทำลายปลายประสาทซิมพาเทติก จะมาพร้อมกับการเพิ่มขึ้นของจำนวนตัวรับเบตาที่ตอบสนองในเนื้อเยื่อ เป็นไปได้ว่าการเพิ่มขึ้นของกิจกรรมของระบบประสาทซิมพาเทติกยังทำให้เกิดการลดความไวต่อสิ่งเร้าที่เกี่ยวข้องกับอายุของหลอดเลือดและเนื้อเยื่อไขมันที่เกี่ยวข้องกับ catecholamine อีกด้วย
จำนวนของตัวรับอะดรีโนในอวัยวะต่างๆ สามารถควบคุมได้ด้วยฮอร์โมนอื่นๆ ดังนั้น เอสตราไดออลจึงเพิ่มขึ้นและโปรเจสเตอโรนจะลดจำนวนของตัวรับอะดรีโนอัลฟาในมดลูก ซึ่งจะมาพร้อมกับการเพิ่มขึ้นและลดลงของการตอบสนองการหดตัวต่อคาเทโคลามีน หาก "ผู้ส่งสารที่สอง" ภายในเซลล์ที่เกิดจากการกระทำของตัวกระตุ้นตัวรับเบต้าเป็น cAMP อย่างแน่นอน สถานการณ์ที่เกี่ยวข้องกับตัวส่งผลของอัลฟา-อะดรีเนอร์จิกจะซับซ้อนมากขึ้น สันนิษฐานว่ามีกลไกต่างๆ อยู่: ระดับของ cAMP ลดลง ปริมาณ cAMP เพิ่มขึ้น การปรับเปลี่ยนไดนามิกของแคลเซียมในเซลล์ เป็นต้น
ในการสร้างผลกระทบต่างๆ ในร่างกาย มักจะต้องใช้อะดรีนาลีนในปริมาณที่น้อยกว่านอร์เอพิเนฟริน 5-10 เท่า แม้ว่านอร์เอพิเนฟรินจะมีประสิทธิภาพมากกว่าเมื่อเปรียบเทียบกับตัวรับอะดรีนาลีนชนิดเอและเบตา 1 แต่ควรจำไว้ว่าคาเทโคลามีนในร่างกายทั้งสองชนิดสามารถโต้ตอบกับตัวรับอัลฟาและเบตาได้ ดังนั้น การตอบสนองทางชีวภาพของอวัยวะใดอวัยวะหนึ่งต่อการกระตุ้นอะดรีเนอร์จิกจึงขึ้นอยู่กับประเภทของตัวรับที่มีอยู่ในอวัยวะนั้นเป็นส่วนใหญ่ อย่างไรก็ตาม นี่ไม่ได้หมายความว่าการกระตุ้นแบบเลือกสรรของระบบประสาทหรือฮิวมอรัลของระบบประสาทซิมพาเทติก-ต่อมหมวกไตเป็นไปไม่ได้ ในกรณีส่วนใหญ่ จะสังเกตเห็นกิจกรรมที่เพิ่มขึ้นของการเชื่อมต่อต่างๆ ของระบบประสาทซิมพาเทติก ดังนั้น จึงยอมรับกันโดยทั่วไปว่าภาวะน้ำตาลในเลือดต่ำจะกระตุ้นต่อมหมวกไตโดยอัตโนมัติ ในขณะที่ความดันโลหิตลดลง (ความดันโลหิตต่ำตามท่าทาง) จะมาพร้อมกับการหลั่งนอร์เอพิเนฟรินจากปลายประสาทซิมพาเทติกเป็นหลัก
ตัวรับต่อมหมวกไตและผลของการกระตุ้นในเนื้อเยื่อต่างๆ
ระบบอวัยวะ |
ชนิดตัวรับอะดรีเนอร์จิก |
ปฏิกิริยา |
ระบบหัวใจและหลอดเลือด: |
||
หัวใจ |
เบต้า |
เพิ่มอัตราการเต้นของหัวใจ การนำไฟฟ้า และการหดตัว |
หลอดเลือดแดงขนาดเล็ก: |
||
ผิวหนังและเยื่อเมือก |
อัลฟ่า |
การลดน้อยลง |
กล้ามเนื้อโครงร่าง |
เบต้า |
การขยายตัว การหดตัว |
อวัยวะในช่องท้อง |
อัลฟา (เพิ่มเติม) |
การลดน้อยลง |
เบต้า |
ส่วนขยาย |
|
เส้นเลือด |
อัลฟ่า |
การลดน้อยลง |
ระบบทางเดินหายใจ: |
||
กล้ามเนื้อของหลอดลม |
เบต้า |
ส่วนขยาย |
ระบบย่อยอาหาร: |
||
ท้อง |
เบต้า |
ทักษะการเคลื่อนไหวลดลง |
ลำไส้ |
อัลฟ่า |
การหดตัวของหูรูด |
ม้าม |
อัลฟ่า |
การลดน้อยลง |
เบต้า |
การผ่อนคลาย |
|
ตับอ่อนมีท่อ |
อัลฟ่า |
การหลั่งลดลง |
ระบบทางเดินปัสสาวะและอวัยวะสืบพันธุ์: |
อัลฟ่า |
การหดตัวของหูรูด |
กระเพาะปัสสาวะ |
เบต้า |
การผ่อนคลายของกล้ามเนื้ออีเจ็คเตอร์ |
อวัยวะเพศชาย |
อัลฟ่า |
การหลั่งน้ำอสุจิ |
ดวงตา |
อัลฟ่า |
การขยายรูม่านตา |
หนัง |
อัลฟ่า |
เหงื่อออกมากขึ้น |
ต่อมน้ำลาย |
อัลฟ่า |
การขับถ่ายโพแทสเซียมและน้ำ |
เบต้า |
การหลั่งอะไมเลส |
|
ต่อมไร้ท่อ: |
||
เกาะของตับอ่อน |
||
เซลล์เบต้า |
อัลฟา (เพิ่มเติม) |
การหลั่งอินซูลินลดลง |
เบต้า |
การหลั่งอินซูลินเพิ่มมากขึ้น |
|
เซลล์อัลฟา |
เบต้า |
เพิ่มการหลั่งกลูคากอน |
8 เซลล์ |
เบต้า |
เพิ่มการหลั่งของโซมาโทสแตติน |
ไฮโปทาลามัสและต่อมใต้สมอง: |
||
โซมาโทโทรฟ |
อัลฟ่า |
เพิ่มการหลั่งของ STH |
เบต้า |
การหลั่ง STH ลดลง |
|
แล็กโทโทรฟ |
อัลฟ่า |
การหลั่งฮอร์โมนโปรแลกตินลดลง |
ไทรอยด์โทรป |
อัลฟ่า |
การหลั่ง TSH ลดลง |
คอร์ติโคโทรป |
อัลฟ่า |
เพิ่มการหลั่ง ACTH |
| เบต้า | การหลั่ง ACTH ลดลง | |
ต่อมไทรอยด์: |
||
เซลล์ฟอลลิคูลาร์ |
อัลฟ่า |
การหลั่งไทรอกซินลดลง |
เบต้า |
เพิ่มการหลั่งของไทรอกซิน |
|
เซลล์พาราฟอลลิคิวลาร์ (K) |
เบต้า |
เพิ่มการหลั่งของแคลซิโทนิน |
ต่อมพาราไทรอยด์ |
เบต้า |
เพิ่มการหลั่งของ PTH |
ไต |
เบต้า |
เพิ่มการหลั่งเรนิน |
ท้อง |
เบต้า |
เพิ่มการหลั่งของแกสตริน |
บีเอ็กซ์ |
เบต้า |
การบริโภคออกซิเจนเพิ่มมากขึ้น |
ตับ |
- |
การเพิ่มขึ้นของการสลายไกลโคเจนและการสร้างกลูโคสใหม่พร้อมกับการปล่อยกลูโคส; การเพิ่มขึ้นของคีโตเจเนซิสพร้อมกับการปล่อยคีโตนบอดี |
เนื้อเยื่อไขมัน |
เบต้า |
เพิ่มการสลายไขมันด้วยการปล่อยกรดไขมันอิสระและกลีเซอรอล |
กล้ามเนื้อโครงร่าง |
เบต้า |
ไกลโคไลซิสเพิ่มขึ้นพร้อมกับการปล่อยไพรูเวตและแลคเตต โปรตีโอไลซิสลดลงพร้อมกับการลดลงของการปล่อยอะลานีน กลูตามีน |
สิ่งสำคัญคือต้องคำนึงว่าผลของการให้ catecholamine ทางเส้นเลือดไม่ได้สะท้อนถึงผลของสารประกอบภายในร่างกายอย่างเพียงพอเสมอไป ซึ่งใช้ได้กับ norepinephrine เป็นหลัก เนื่องจากในร่างกาย norepinephrine จะถูกปล่อยออกมาโดยตรงที่ช่องซินแนปส์ ไม่ใช่ในเลือด ดังนั้น norepinephrine ในร่างกายจึงไม่เพียงแต่กระตุ้นตัวรับอัลฟาในหลอดเลือด (ทำให้ความดันโลหิตสูงขึ้น) เท่านั้น แต่ยังกระตุ้นตัวรับเบตาของหัวใจ (ทำให้หัวใจเต้นเร็วขึ้น) ด้วย ในขณะที่การได้รับ norepinephrine จากภายนอกจะกระตุ้นตัวรับอัลฟาในหลอดเลือดเป็นหลัก และการเต้นของหัวใจช้าลงโดยรีเฟล็กซ์ (ผ่านเส้นประสาทเวกัส)
อะดรีนาลีนในปริมาณต่ำจะไปกระตุ้นตัวรับเบต้าของหลอดเลือดกล้ามเนื้อและหัวใจเป็นหลัก ส่งผลให้ความต้านทานของหลอดเลือดส่วนปลายลดลงและปริมาณเลือดที่สูบฉีดออกจากหัวใจเพิ่มขึ้น ในบางกรณี อาจเกิดผลแรก และความดันโลหิตต่ำจะเกิดขึ้นหลังจากใช้อะดรีนาลีน ในปริมาณที่สูงขึ้น อะดรีนาลีนจะไปกระตุ้นตัวรับอัลฟาด้วย ซึ่งจะมาพร้อมกับความต้านทานของหลอดเลือดส่วนปลายที่เพิ่มขึ้น และเมื่อปริมาณเลือดที่สูบฉีดออกจากหัวใจเพิ่มขึ้น จะทำให้ความดันเลือดแดงเพิ่มขึ้น อย่างไรก็ตาม ผลของอะดรีนาลีนต่อตัวรับเบต้าของหลอดเลือดก็ยังคงมีอยู่เช่นกัน ส่งผลให้ความดันซิสโตลิกเพิ่มขึ้นเกินกว่าตัวบ่งชี้ความดันไดแอสโตลิกที่คล้ายกัน (ความดันชีพจรเพิ่มขึ้น) เมื่อใช้ยาในปริมาณที่สูงขึ้น ผลที่เลียนแบบอัลฟาของอะดรีนาลีนจะเริ่มมีมากขึ้น โดยความดันซิสโตลิกและไดแอสโตลิกจะเพิ่มขึ้นควบคู่กัน เช่นเดียวกับภายใต้อิทธิพลของนอร์เอพิเนฟริน
ผลกระทบของคาเทโคลามีนต่อการเผาผลาญประกอบด้วยผลโดยตรงและโดยอ้อม โดยผลโดยตรงเกิดขึ้นจากตัวรับเบตาเป็นส่วนใหญ่ กระบวนการที่ซับซ้อนกว่านั้นเกี่ยวข้องกับตับ แม้ว่าโดยทั่วไปแล้วการสลายไกลโคเจนของตับที่เพิ่มขึ้นจะถือได้ว่าเป็นผลจากการกระตุ้นตัวรับเบตา แต่ก็มีหลักฐานว่าตัวรับอัลฟามีส่วนเกี่ยวข้องด้วย ผลทางอ้อมของคาเทโคลามีนสัมพันธ์กับการปรับการหลั่งของฮอร์โมนอื่นๆ มากมาย เช่น อินซูลิน ในผลของอะดรีนาลีนต่อการหลั่งนั้น ส่วนประกอบอัลฟาอะดรีเนอร์จิกมีอิทธิพลเหนือกว่าอย่างชัดเจน เนื่องจากมีการแสดงให้เห็นว่าความเครียดใดๆ จะมาพร้อมกับการยับยั้งการหลั่งอินซูลิน
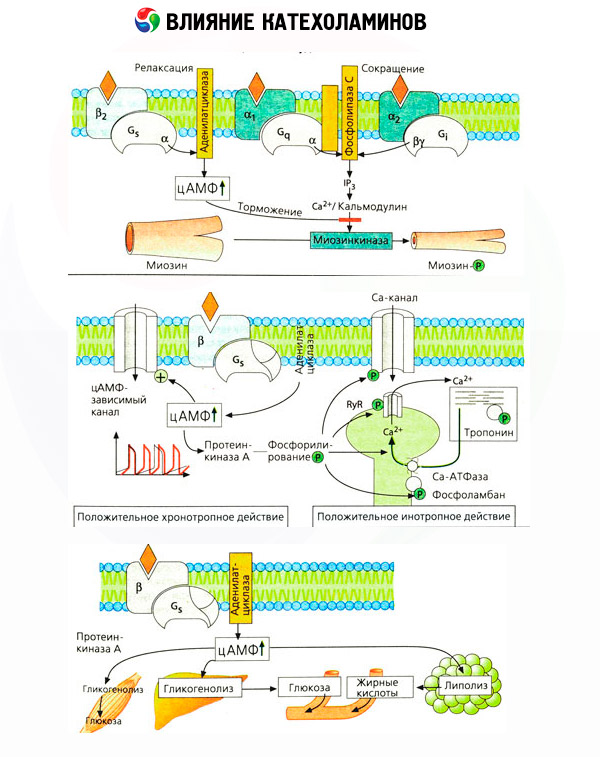
การรวมกันของผลทางตรงและทางอ้อมของ catecholamine ทำให้เกิดภาวะน้ำตาลในเลือดสูง ซึ่งเกี่ยวข้องไม่เพียงแต่กับการผลิตกลูโคสในตับที่เพิ่มขึ้นเท่านั้น แต่ยังรวมถึงการยับยั้งการใช้กลูโคสในเนื้อเยื่อรอบนอกอีกด้วย การเร่งการสลายไขมันทำให้เกิดภาวะไขมันในเลือดสูง ซึ่งส่งผลให้มีการส่งกรดไขมันไปที่ตับมากขึ้น และการผลิตคีโตนบอดีเพิ่มขึ้น ไกลโคไลซิสในกล้ามเนื้อที่เพิ่มขึ้นส่งผลให้มีการปลดปล่อยแลคเตตและไพรูเวตเข้าสู่เลือดมากขึ้น ซึ่งเมื่อรวมกับกลีเซอรอลที่ปลดปล่อยจากเนื้อเยื่อไขมัน จะทำหน้าที่เป็นสารตั้งต้นของกระบวนการสร้างกลูโคสใหม่ในตับ
การควบคุมการหลั่งคาเทโคลามีน ความคล้ายคลึงกันของผลิตภัณฑ์และวิธีการตอบสนองของระบบประสาทซิมพาเทติกและต่อมหมวกไตส่วนในเป็นพื้นฐานสำหรับการรวมโครงสร้างเหล่านี้เข้าเป็นระบบซิมพาเทติก-ต่อมหมวกไตเดียวของร่างกายพร้อมการจัดสรรการเชื่อมโยงประสาทและฮอร์โมน สัญญาณรับเข้าต่างๆ จะรวมตัวอยู่ในไฮโปทาลามัสและศูนย์กลางของไขสันหลังและเมดัลลาอ็อบลองกาตา ซึ่งเป็นจุดที่ข้อความส่งออกเริ่มต้นขึ้น โดยสลับไปที่เซลล์ของนิวรอนก่อนปมประสาทที่ตั้งอยู่ในส่วนด้านข้างของไขสันหลังในระดับของส่วนคอ VIII - ส่วนเอว II-III
แอกซอนก่อนปมประสาทของเซลล์เหล่านี้จะออกจากไขสันหลังและสร้างการเชื่อมต่อแบบซินแนปส์กับเซลล์ประสาทที่อยู่ในปมประสาทของห่วงโซ่ซิมพาเทติกหรือกับเซลล์ของต่อมหมวกไตส่วนใน เส้นใยก่อนปมประสาทเหล่านี้เป็นโคลีเนอร์จิก ความแตกต่างพื้นฐานประการแรกระหว่างเซลล์ประสาทหลังปมประสาทซิมพาเทติกและเซลล์โครมาฟฟินของต่อมหมวกไตส่วนในก็คือ เซลล์โครมาฟฟินของต่อมหมวกไตส่วนในส่งสัญญาณโคลีเนอร์จิกที่รับโดยเซลล์เหล่านี้ไม่ใช่ผ่านการนำสัญญาณของเส้นประสาท (เส้นประสาทอะดรีเนอร์จิกหลังปมประสาท) แต่ผ่านเส้นทางฮิวมอรัล โดยปล่อยสารประกอบอะดรีเนอร์จิกเข้าสู่กระแสเลือด ความแตกต่างประการที่สองก็คือ เส้นประสาทหลังปมประสาทสร้างนอร์เอพิเนฟริน ในขณะที่เซลล์ของต่อมหมวกไตส่วนในจะผลิตอะดรีนาลีนเป็นหลัก สารทั้งสองนี้มีผลต่อเนื้อเยื่อต่างกัน