ผู้เชี่ยวชาญทางการแพทย์ของบทความ

สิ่งตีพิมพ์ใหม่
รังไข่
ตรวจสอบล่าสุด: 23.04.2024

เนื้อหา iLive ทั้งหมดได้รับการตรวจสอบทางการแพทย์หรือตรวจสอบข้อเท็จจริงเพื่อให้แน่ใจว่ามีความถูกต้องตามจริงมากที่สุดเท่าที่จะเป็นไปได้
เรามีแนวทางการจัดหาที่เข้มงวดและมีการเชื่อมโยงไปยังเว็บไซต์สื่อที่มีชื่อเสียงสถาบันการวิจัยทางวิชาการและเมื่อใดก็ตามที่เป็นไปได้ โปรดทราบว่าตัวเลขในวงเล็บ ([1], [2], ฯลฯ ) เป็นลิงก์ที่คลิกได้เพื่อการศึกษาเหล่านี้
หากคุณรู้สึกว่าเนื้อหาใด ๆ ของเราไม่ถูกต้องล้าสมัยหรือมีข้อสงสัยอื่น ๆ โปรดเลือกแล้วกด Ctrl + Enter
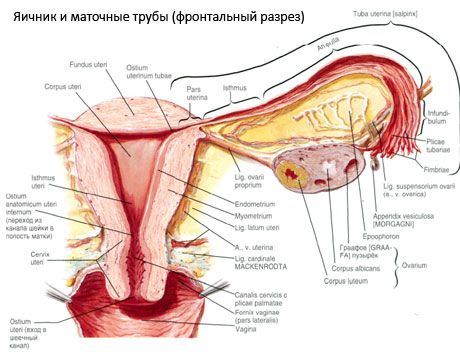
รังไข่ (ovarium, oophoron ของชาวกรีก) - อวัยวะที่จับคู่อวัยวะเพศหญิงตั้งอยู่ในโพรงของกระดูกเชิงกรานขนาดเล็กที่อยู่หลังเอ็นกว้างของมดลูก รังไข่พัฒนาและพัฒนาเซลล์เพศหญิง (ovules) รวมทั้งฮอร์โมนเพศหญิงที่เข้าสู่กระแสเลือดและน้ำเหลือง รังไข่มีรูปร่างเป็นวงรีค่อนข้างแบนในทิศทางก่อนหน้า สีของรังไข่มีสีชมพู บนพื้นผิวของรังไข่ของหญิงที่ให้กำเนิดความหดหู่และรอยแผลเป็นสามารถมองเห็นได้ - ร่องรอยของการตกไข่และการเปลี่ยนสีเหลือง รังไข่มีขนาด 5-8 กรัมรังไข่มีขนาด 2.5-5.5 ซม. ยาว 1.5-3.0 ซม. กว้าง 2 เซนติเมตรรังไข่มีสองชั้นฟรี ) หันไปทางช่องเชิงกรานปกคลุมบางส่วนท่อนำไข่และพื้นผิวด้านข้าง (facies lateralis) ติดผนังด้านข้างกระดูกเชิงกรานลึกอ่อน - แอ่งรังไข่ หลุมฝังศพนี้ตั้งอยู่ที่มุมระหว่างช่องท้องด้านนอกของช่องท้องด้านบนและด้านหลังของหลอดเลือดแดงมดลูกและลำไส้เล็กส่วนล่าง หลังรังไข่, ท่อปัสสาวะที่ด้านข้างตรงกันข้ามจะผ่านจากบนลงล่าง
พื้นผิวรังไข่ไปในนูนฟรี (หลัง) ขอบ (Margo Liber) หน้า - ในภูมิภาค mesenteric (Margo mesovaricus) โดยใช้วิธีการระยะสั้นคลิปพับของเยื่อบุช่องท้อง (น้ำเหลืองรังไข่) กับใบหลังของเอ็นกว้างของมดลูก อยู่ในระดับแนวหน้าของร่างกายเป็นร่องพักผ่อน - ประตูรังไข่ (หลุม ovarii) ผ่านที่รังไข่รวมถึงเส้นเลือดและเส้นประสาทเส้นเลือดและตั้งอยู่ lymphatics นอกจากนี้เรายังแยกรังไข่ทั้งสองปลาย: กลมบนปลายท่อ (extremitas tubaria) หันหน้าไปทางท่อนำไข่และปลายล่างของแม่ (extremitas utenna) ควบคู่กับมดลูกพวงเองรังไข่ (lig ovarii proprium.) มัดในสายรอบประมาณ 6 มมจากแม่นี่คือจุดสิ้นสุดไปยังด้านข้างมุมมดลูกรังไข่อยู่ระหว่างสองแผ่นเอ็นกว้าง โดยรังไข่อุปกรณ์เอ็นยังเกี่ยวข้องเอ็น podveshivayaschaya รังไข่ (lig.suspensorium ovarii) ซึ่งเป็นพับของเยื่อบุช่องท้องยื่นออกมาจากผนังด้านบนของกระดูกเชิงกรานเพื่อรังไข่และรังไข่ที่มีเรือและการรวมกลุ่มเส้นใยของเส้นใย รังไข่คงสั้นน้ำเหลือง (mesovarium) ซึ่งหมายถึงเยื่อบุช่องท้อง duplikatury ขยายจากเอ็นกว้างหลังใบปลิวของมดลูกไปที่ขอบ mesenteric รังไข่ รังไข่ตัวเองไม่ได้รับการปกคลุมด้วย peritoneum ในตอนท้ายของหลอดที่แนบมารังไข่ fimbria รังไข่ขนาดใหญ่ที่สุดของท่อนำไข่ ภูมิประเทศของรังไข่ขึ้นอยู่กับตำแหน่งของมดลูก, ความคุ้มค่า (ในการตั้งครรภ์) รังไข่หมายถึงอวัยวะที่เคลื่อนที่มากในโพรงในอุ้งเชิงกราน
 [
[หลอดเลือดและเส้นประสาทของรังไข่
การจัดหาเลือดไปยังรังไข่เกิดจาก aa. และอื่น ๆ ovaricae et uterinae (. Aa ovaricae Dextra et sinistra) ทั้งหลอดเลือดแดงรังไข่เพิ่มขึ้นจากพื้นผิวด้านหน้าของหลอดเลือดแดงใหญ่อยู่ด้านล่างของหลอดเลือดแดงไตขวาที่ส่วนใหญ่มักจะมาจากหลอดเลือดแดงใหญ่และซ้าย - ของหลอดเลือดแดงไต จะลงและด้านข้างมากกว่ากล้ามเนื้อหน้า psoas ด้านหน้าหลอดเลือดรังไข่ปริภูมิแต่ละท่อไตด้านหน้า (ให้เขากิ่งไม้), เรืออุ้งเชิงกรานภายนอกและเส้นเขตแดนเข้ามาในโพรงกระดูกเชิงกรานถูกตั้งอยู่ที่นี่ในเอ็นแขวนของรังไข่ ต่อไปนี้ตรงกลางหลอดเลือดรังไข่ผ่านระหว่างแผ่นเอ็นกว้างของมดลูกภายใต้ท่อนำไข่ให้มันสาขาและจากนั้น - ในน้ำเหลืองของรังไข่; เข้าประตูรั้วรังไข่
สาขาของหลอดเลือดแดงที่เกี่ยวกับรังไข่มีส่วนเกี่ยวข้องกับแขนงที่เกี่ยวกับรังไข่ของหลอดเลือดแดงมดลูก การไหลเวียนของรังไข่จากรังไข่จะกระทำโดยหลักในรังไข่เยื่อหุ้มปอดซึ่งตั้งอยู่ในบริเวณประตูรังไข่ ดังนั้นการไหลออกของเลือดจะผ่านไปสองทิศทาง: ผ่านทางหลอดเลือดดำมดลูกและรังไข่ หลอดเลือดดำด้านขวาของหลอดเลือดดำมีวาล์วและวิ่งเข้าไปใน vena cava ที่ต่ำกว่า เส้นเลือดบีบซ้ายจะไหลเข้าสู่หลอดเลือดดำด้านซ้ายโดยไม่มีวาล์วอยู่
Lymph outflow จากรังไข่เกิดขึ้นผ่านทางเรือน้ำเหลืองโดยเฉพาะอย่างยิ่งในพื้นที่ของประตูอวัยวะที่ plexus น้ำเหลืองใต้ลิ้นจะถูกขับออกมา จากนั้นต่อมน้ำเหลืองจะถูกนำไปยังต่อมน้ำเหลืองพาราพลาสมาไปตามลำไส้เล็กของรังไข่
Innervation ของรังไข่
เห็นใจ - ให้เส้นใยจากเซลล์ celag (แสงอาทิตย์) postganglionic, plexuses บนถักและ hypogastric; parasympathetic - เนื่องจากเส้นประสาทภายใน sacral
โครงสร้างของรังไข่
พื้นผิวที่ถูกปกคลุมด้วยชั้นเดียวของเยื่อบุผิวเชื้อโรครังไข่ ใต้มันโกหก albuginea เกี่ยวพันหนาแน่น (กอช albuginea) เนื้อเยื่อเกี่ยวพันในรูปแบบที่รังไข่ stroma (stroma ovarii) ที่อุดมไปด้วยเส้นใยที่มีความยืดหยุ่น สารเนื้อเยื่อรังไข่มันโดยแบ่งเป็นชั้นนอกและด้านใน ชั้นในซึ่งตั้งอยู่ในใจกลางของรังไข่ที่ใกล้ชิดกับประตูที่เรียกว่าไขกระดูก (ไขกระดูก ovarii) ในชั้นนี้ในเนื้อเยื่อเกี่ยวพันหลวมเป็นจำนวนมากในเลือดและน้ำเหลืองเรือและเส้นประสาท ชั้นนอกของรังไข่ - เปลือกนอก ovarii มีความหนาแน่นมากขึ้น มันมีจำนวนมากของเนื้อเยื่อเกี่ยวพันในการที่จะอยู่สุกรังไข่หลัก (folliculi ovarici primarii) รอง (ฟอง) รูขุมขน (folliculi ovarici secundarii, s.vesiculosi) ดีรูขุมผู้ใหญ่ graafovy ฟอง (folliculi ovarici maturis) และสีเหลือง และร่างกาย atretic
ในรูขุมขนแต่ละเซลล์จะเป็นไข่เพศเมียหรือ oocyte (ovocytus) เส้นผ่าศูนย์กลางไข่ได้ถึง 150 ไมครอนกลมมีนิวเคลียสจำนวนมากของพลาสซึมซึ่งนอกเหนือไปจาก organelles โทรศัพท์มือถือมีการรวมโปรตีนไขมัน (ไข่แดง) ไกลโคเจนที่จำเป็นสำหรับการจัดหาไข่ ปริมาณไข่จะกินภายใน 12-24 ชั่วโมงหลังการตกไข่ ถ้าการปฏิสนธิไม่เกิดขึ้นไข่จะตาย
รังไข่ของมนุษย์มีเยื่อหุ้มบัง 2 ข้าง ภายในเป็นcytolemmaซึ่งเป็นเยื่อ cytoplasmic ของ oocyte ด้านนอก cytolemma มีชั้นของเซลล์ follicular ที่เรียกว่าปกป้องไข่และทำหน้าที่สร้างฮอร์โมน - พวกเขาปล่อย estrogens
ตำแหน่งทางสรีรวิทยาของโพรงมดลูกหลอดและรังไข่มีให้โดยเครื่องระงับการยึดติดและอุปกรณ์เสริมประกอบกับเยื่อบุช่องท้องเอ็นและเซลลูโลสในอุ้งเชิงกราน อุปกรณ์ระงับจะแสดงโดยการก่อตัวคู่ซึ่งจะรวมถึงเส้นเอ็นรอบและกว้างของมดลูกเอ็นของตัวเองและที่แขวนอยู่ด้านข้างรังไข่ เอ็นกว้างของมดลูกเอ็นเอ็นและแขวนลอยของรังไข่ทำให้มดลูกอยู่ในตำแหน่งกลาง เส้นเอ็นรอบด้านดึงดูดส่วนล่างของมดลูกไว้ข้างหน้าและให้ความเอียงทางสรีรวิทยา
อุปกรณ์ยึดยึดตำแหน่งของตัวสั่นในศูนย์กลางของกระดูกเชิงกรานขนาดเล็กและทำให้แทบจะเป็นไปไม่ได้ที่จะเลื่อนไปทางด้านข้างไปมา แต่เป็นเส้นเอ็นของมดลูกย้ายจากส่วนล่างของความชอบที่เป็นไปได้ของมดลูกในทิศทางที่แตกต่างกัน โดยกำหนดหน่วยรวมถึงสายจัดในเนื้อเยื่อกระดูกเชิงกรานหลวมและยื่นออกมาจากการ์ดมดลูกลดลงไปที่ด้านข้างด้านหน้าและผนังด้านหลังของกระดูกเชิงกราน: sacroiliac magochnye พระคาร์ดินัลมดลูกและเอ็นเปาะ-vesico-pubic
นอกเหนือไปจาก mesovarium เอ็นสตรีรังไข่มีลักษณะเด่นดังนี้
- เอ็นแขวนของรังไข่กำหนดก่อนหน้านี้เป็น voronkotazovaya มันเป็นพับของเยื่อบุช่องท้องด้วยการขยายนั้นเลือด (ก. Et v. Ovarica) และท่อน้ำเหลืองและเส้นประสาทรังไข่ยืดระหว่างผนังกับกระดูกเชิงกรานด้านข้างป้ายเอว (ในหารหลอดเลือดอุ้งเชิงกรานร่วมกันที่ด้านนอกและด้าน) และบน (หลอด) ปลายรังไข่;
- เอ็นของเอ็นรังไข่ผ่านระหว่างเอ็นของเอ็นมดลูกกว้างใกล้กับแผ่นด้านหลังและเชื่อมต่อด้านล่างของรังไข่กับด้านข้างของมดลูก กับมดลูกเอ็นเอ็นของตัวเองที่ติดอยู่ระหว่างจุดเริ่มต้นของท่อมดลูกกับเอ็นที่เป็นวงกลมไปมาจากหลัง ในความหนาของเอ็นคือ rr. Ovarii ซึ่งเป็นสาขาปลายของเส้นเลือดแดงมดลูก
- ลำไส้รังไข่ยื่นออกมาเกี่ยวกับสันเขาด้านนอกของรังไข่ด้านขวาหรือเอ็นกว้างของมดลูกในรูปแบบของรอยพับของเยื่อบุโพรงมดลูก เอ็นเป็นที่ไม่เสถียรและสังเกตเห็นได้ในผู้หญิงที่เป็น 1/2 ถึง 1/3
เครื่องช่วยแสดงโดยกล้ามเนื้อและพังผืดของอุ้งเชิงกรานแบ่งออกเป็นชั้นล่างกลางและบน (ภายใน)
ที่มีประสิทธิภาพมากที่สุดคือชั้นกล้ามเนื้อด้านบน (ด้านใน) ซึ่งแสดงโดยกล้ามเนื้อจับคู่ที่ยกทวารหนัก ประกอบด้วยกล้ามเนื้อมัดที่พัดออกจากกระดูกก้นกบไปยังกระดูกเชิงกรานในสามทิศทาง (pubic-coccygeal, ilio-coccygeal และ ischiococcycyalal กล้ามเนื้อ) ชั้นของกล้ามเนื้อนี้เรียกว่าไดอะแฟรมของกระดูกเชิงกราน
ชั้นกลางของกล้ามเนื้อตั้งอยู่ระหว่างกระดูกพยุงกระดูกตีบและกระดูก ischium ชั้นกลางของกล้ามเนื้อ - ไดอะแฟรมของระบบทางเดินปัสสาวะ - ใช้ครึ่งทางก่อนหน้าของช่องท้องอุ้งเชิงกรานโดยผ่านทางท่อปัสสาวะและช่องคลอด ในส่วนหน้าระหว่างใบของมันเป็นคานกล้ามเนื้อสร้างกล้ามเนื้อหูรูดด้านนอกของท่อปัสสาวะในส่วนหลังมีการรวมกลุ่มของกล้ามเนื้อไปในทิศทางขวางกล้ามเนื้อขวางลึกของ perineum
ที่ต่ำกว่า (นอก) ชั้นของกล้ามเนื้ออุ้งเชิงกรานประกอบด้วยพื้นผิวซึ่งสถานที่มีรูปร่างเหมือนรูปที่ 8 เหล่านี้รวมถึง bulbospongiosus-โพรง ischio-โพรงกล้ามเนื้อหูรูดของทวารหนักภายนอกที่ตื้นกล้ามเนื้อฝีเย็บขวาง
วิวัฒนาการของรังไข่
กระบวนการของการเจริญเติบโตและ atresia follicular เริ่มต้นด้วยการตั้งครรภ์ 20 สัปดาห์และตามเวลาของการจัดส่งในรังไข่ของหญิงสาวยังคงได้ถึง 2 ล้านเซลล์ไข่ เมื่อถึงวัยหมดประจำเดือนจำนวนของพวกเขาลดลงเป็น 300,000 ในช่วงระยะเวลาทั้งหมดของชีวิตการเจริญพันธุ์ถึงวุฒิภาวะและไม่เกิน 500 รูขุมขน การเจริญเติบโตครั้งแรกของรูขุมขนไม่ได้ขึ้นอยู่กับการกระตุ้นของ FSH มีข้อ จำกัด และอาการ atresia เกิดขึ้นได้อย่างรวดเร็ว เป็นที่เชื่อกันว่าในตำแหน่งของเตียรอยด์ฮอร์โมนเปปไทด์ autocrine ท้องถิ่น / paracrine เป็นตัวควบคุมหลักของการเจริญเติบโตและ atresia ของรูขุมขนหลัก เป็นที่เชื่อกันว่ากระบวนการของการเจริญเติบโตและ atresia ของรูขุมไม่ขัดจังหวะโดยกระบวนการทางสรีรวิทยาใด ๆ กระบวนการนี้ยังคงดำเนินต่อไปในทุกช่วงอายุรวมถึงช่วงตั้งครรภ์และวัยหมดประจำเดือนซึ่งถูกขัดจังหวะด้วยการตั้งครรภ์การตกไข่และการเกิด anovulation กลไกที่ทำให้เกิดการเจริญเติบโตของรูขุมขนและจำนวนของพวกเขาในแต่ละวัฏจักรที่เฉพาะเจาะจงยังไม่ชัดเจน
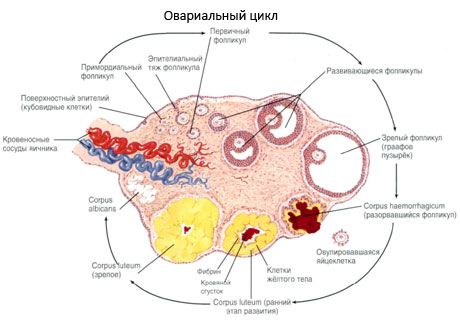
ในการพัฒนารูขุมขนผ่านขั้นตอนต่างๆของการพัฒนา เซลล์สืบเชื้อสายมาจาก endoderm ของถุงไข่แดง allantois และโยกย้ายไปยังบริเวณอวัยวะเพศของตัวอ่อนในสัปดาห์ที่ 5-6 ของการตั้งครรภ์ อันเป็นผลมาจากการผ่าเหล่าอย่างรวดเร็วซึ่งมีระยะเวลาตั้งแต่ 6-8 สัปดาห์ถึง 16-20 สัปดาห์ของการตั้งครรภ์จะมีเซลล์ไข่ถึงประมาณ 6-7 ล้านเซลล์ในรังไข่ซึ่งล้อมรอบด้วยเซลล์เม็ดเล็ก ๆ บาง ๆ
รูขุมขนก่อนวัย - ไข่ล้อมรอบด้วยเยื่อหุ้มปอด (Zona pellucida) เซลลูโลสเซลล์รอบไข่เริ่มเจริญเติบโตการเจริญเติบโตของพวกเขาขึ้นอยู่กับ gonadotropins และมีความสัมพันธ์กับระดับของ estrogens Granulosa cells เป็นเป้าหมายของ FSH ในขั้นตอนของรูขุมขนก่อนเซลล์เม็ดโลหิตสามารถสังเคราะห์สเตียรอยด์ได้สามชั้น: กระตุ้นให้เกิดกิจกรรมของ aromatase ซึ่งเป็นเอนไซม์หลักที่แปลง androgens เป็น estradiol มีความเชื่อกันว่า estradiol สามารถเพิ่มจำนวนผู้รับของตัวเองได้ทำให้มีผลต่อ mitogenic โดยตรงต่อเซลล์ granulosa ที่ไม่ขึ้นกับ FSH ถือเป็นปัจจัยของ paracrine ที่ช่วยเพิ่มผลกระทบของ FSH รวมถึงการกระตุ้นกระบวนการ aromatization
ตัวรับ FSH จะปรากฏบนเยื่อหุ้มเซลล์ของ granulosa เมื่อการเจริญเติบโตของรูขุมขนเริ่มขึ้น การลดหรือการเพิ่มขึ้นของ FSH ทำให้เกิดการเปลี่ยนแปลงจำนวนผู้รับ การกระทำของ FSH นี้ถูกปรับโดยปัจจัยการเจริญเติบโต FSH ทำหน้าที่ผ่าน G-โปรตีนระบบ adenylate cyclase-ในรูขุมขน steroidogenesis แม้ว่าส่วนใหญ่ที่มีการควบคุม FSH กระบวนการที่เกี่ยวข้องกับปัจจัยหลายช่องไอออนระบบรับซายน์ไคเนส phospholipase ทูตรอง
บทบาทของ androgens ในการพัฒนาต้นของรูขุมขนนั้นมีความซับซ้อน เซลล์ Granulosa มีตัวรับแอนโดรเจน พวกเขาจะไม่เพียง แต่สารตั้งต้นสำหรับ aromatization FSH ที่เกิดขึ้นลงในสโตรเจน แต่ในความเข้มข้นต่ำที่สามารถเพิ่มประสิทธิภาพกระบวนการของเครื่องปรุง เมื่อระดับของแอนโดรเจนที่เพิ่มขึ้นเซลล์ preantral granulosa เลือกพิเศษเส้นทาง aromatization ไม่มีเข้า estrogens และวิธีที่ง่ายเพื่อการแปลงผ่าน androgens 5A-reductase ในแอนโดรเจนพัฒนาที่ไม่สามารถแปลงเป็นสโตรเจนและกิจกรรม aromatase จึงยับยั้ง กระบวนการนี้ยังยับยั้งการก่อตัวของ FSH และ LH ผู้รับจึงหยุดการพัฒนาของรูขุมขน
กระบวนการ aromatization, follicle ที่มีระดับสูงของ androgens ผ่านกระบวนการ atresia การเจริญเติบโตและการพัฒนาของรูขุมขนขึ้นอยู่กับความสามารถในการแปลง androgens เป็น estrogens
ในการปรากฏตัวของ FSH สารที่เด่นชัดของน้ำนมคือ folrogens ในกรณีที่ไม่มี FSH - androgens LH เป็นปกติใน fluid follicular จนถึงกลางของรอบ ทันทีที่ระดับที่เพิ่มขึ้นของกิจกรรมทิคส์ของเซลล์ granulosa LH ลดลงและการเปลี่ยนแปลงเสื่อมปรากฏเพิ่มระดับแอนโดรเจนในระดับรูขุมขนเตียรอยด์ในของเหลว follicular กว่าในพลาสมาและสะท้อนให้เห็นถึงการทำงานของรังไข่กิจกรรม: granulosa และ theca เซลล์ ถ้าเป้าหมายเพียงอย่างเดียวสำหรับ FSH คือเซลล์ granulosa แล้ว LH มีเป้าหมายมากมายซึ่ง ได้แก่ เซลล์ stromal และ luteal cells และ granulosa cells ความสามารถในการ steroidogenesis มีทั้ง granulosa และ teka เซลล์ แต่กิจกรรม aromatase dominant ในเซลล์ granulosa.
ในการตอบสนองต่อ LH เซลล์ Teka ผลิต androgens ซึ่งผ่านทาง aromatization ที่เกิดจาก FSH จะถูกเปลี่ยนจากเซลล์ granulosa ไปเป็น estrogens
ในฐานะที่เป็นรูขุมขนเซลล์ theca เริ่มต้นที่จะแสดงยีนสำหรับ LH รับ P450 วินาทีและ 3beta-hydroxysteroid dehydrogenase, อินซูลินเช่นปัจจัยการเจริญเติบโต (IGF-1) ร่วมกับ LH เพื่อเพิ่มการแสดงออกของยีน แต่ไม่ได้ช่วยกระตุ้น steroidogenesis
การเตียรอยด์ของรังไข่อยู่เสมอขึ้นอยู่กับ LH ในขณะที่รูขุมขนเติบโตขึ้นเซลล์ในปัจจุบันจะแสดงเอนไซม์ P450c17 ซึ่งทำหน้าที่เป็นแอนโดรเจนจากคอเลสเตอรอล เซลล์ Granulosa ไม่ได้มีเอนไซม์นี้และขึ้นอยู่กับเซลล์ปัจจุบันในการผลิตฮอร์โมนเอสโตรเจนจาก androgens แตกต่างจาก steroidogenesis - folliculogenesis ขึ้นอยู่กับ FSH ในฐานะที่เป็นรูขุมขนและเพิ่มระดับของสโตรเจนที่เข้ามาในกลไกความคิดเห็นการดำเนินการ - การผลิตยับยั้งของ FSH ซึ่งจะนำไปสู่การลดลงในรูขุมขนกิจกรรม aromatase และท้ายที่สุดจะตีบตันรูขุมขนผ่าน apoptosis (โปรแกรมมือถือตาย)
กลไกการตอบสนองของฮอร์โมนเอสโตรเจนและ FSH ยับยั้งการพัฒนารูขุมขนที่เริ่มมีการเจริญเติบโต แต่ไม่ใช่รูขุมขนที่สำคัญ รูขุมขนที่เด่น ๆ มีตัวรับ FSH มากขึ้นที่สนับสนุนการงอกของเซลล์เม็ดเลือดและการทำ aromatization ของ androgens ใน estrogens นอกจากนี้ paracrine และ autocrine pathway ยังทำหน้าที่เป็นผู้ประสานงานที่สำคัญสำหรับการพัฒนารูขุมขน antral follicle
ส่วนประกอบสำคัญของสารควบคุม autocrine / paracrine คือเปปไทด์ (inhibin, activin, follistatin) ซึ่งสังเคราะห์ขึ้นโดยเซลล์ granulosa เพื่อตอบสนองต่อการกระทำของ FSH และเข้าสู่ follicular fluid Inhibin ช่วยลดการหลั่ง FSH activin กระตุ้นการปลดปล่อย FSH จากต่อมใต้สมองและช่วยเพิ่มการทำงานของ FSH ในรังไข่ Follistatin ยับยั้งกิจกรรม FSH อาจเป็นเพราะการมีส่วนร่วมของ activin หลังจากการตกไข่และการพัฒนาตัวเหลือง inhibin อยู่ภายใต้การควบคุมของ LH
การเจริญเติบโตและความแตกต่างของเซลล์รังไข่ได้รับอิทธิพลจากปัจจัยการเติบโตของอินซูลิน (IGE) IGF-1 ทำหน้าที่ในเซลล์ granulosa ทำให้เกิด cyclic adenosine monophosphate (cAMP), progesterone, oxytocin, proteoglycan และ inhibin
IGF-1 ทำหน้าที่ในเซลล์ teka ทำให้เกิดการเพิ่มขึ้นของ androgens เซลล์ Teka ทำให้เกิดปัจจัยการตายของเนื้อร้ายเนื้องอก (TNF) และปัจจัยการเจริญเติบโตของผิวหนัง (epidermal growth factor - EGF) ซึ่งควบคุมโดย FSH
EGF ช่วยกระตุ้นการขยายตัวของเซลล์เม็ดเลือด IGF-2 เป็นปัจจัยหลักในการเจริญเติบโตของเชื้อ follicular fluid นอกจากนี้ยังตรวจพบ IGF-1, TNF-a, TNF-3 และ EGF
การละเมิดกฎของ paracrine และ / หรือ autocrine ในการทำงานของรังไข่ดูเหมือนจะมีบทบาทในการรบกวนกระบวนการตกไข่และในการสร้างรังไข่ polycystic
ในฐานะที่เป็นรูขุมขนเติบโตขึ้นเนื้อหาของ estrogens ใน follicular fluid เพิ่มขึ้น ที่จุดสูงสุดของการเพิ่มขึ้นของพวกเขาในเซลล์ granulosa, ผู้รับสำหรับ LH ปรากฏ luteinization ของเซลล์ granulosa เกิดขึ้นและการผลิต progesterone เพิ่มขึ้น ดังนั้นในช่วงก่อนการทดลองการเพิ่มขึ้นของการผลิตฮอร์โมนเอสโตรเจนทำให้เกิดตัวรับ LH LH จึงทำให้เกิด luteinization ของเซลล์เม็ดเลือดและการผลิต progesterone การเพิ่มขึ้นของฮอร์โมนโปรเจสเตอโรนทำให้ระดับฮอร์โมนเอสโตรเจนลดลงซึ่งเห็นได้ชัดว่าเป็นสาเหตุให้เกิด FSH ที่สองในช่วงกลางของวัฏจักร
เป็นที่เชื่อกันว่าการตกไข่เกิดขึ้น 10-12 ชั่วโมงหลังจากที่จุดสูงสุดของ LH และ 24-36 ชั่วโมงหลังจากจุดสูงสุดของ estradiol เป็นที่เชื่อกันว่า LH ช่วยกระตุ้นการลด oocyte, luteinization ของเซลล์ granulosa, การสังเคราะห์ progesterone และ prostaglandin ใน follicle
โปรเจสเตอโรนช่วยเพิ่มเอนไซม์โปรตีเอสโตริกรวมทั้ง prostaglandin ที่เกี่ยวข้องกับการแตกของรูขุมขน FSH เหนี่ยวนำให้เกิดยอดกระเทือนช่วยให้การส่งออกของไข่จากรูขุมขนโดยการแปลง plasminogen กับเอนไซม์ - plasmin ให้ในปริมาณที่เพียงพอของตัวรับ LH สำหรับการพัฒนาปกติของเฟส luteal
ภายใน 3 วันหลังจากการตกไข่ granulosa เซลล์เพิ่มขึ้นของพวกเขาปรากฏ vacuoles ลักษณะที่เต็มไปด้วยเม็ดสี - ลูทีน เซลล์ theca-ลูทีนแตกต่างจาก theca และ stroma และกลายเป็นส่วนหนึ่งของ luteum คลัง ได้อย่างรวดเร็วภายใต้อิทธิพลของปัจจัย angiogenic คือการพัฒนาของเส้นเลือดฝอยที่ซึม luteum คลังและเพื่อปรับปรุง vascularization เพิ่มการผลิตฮอร์โมนและสโตรเจน steroidogenesis กิจกรรมและระยะเวลาของชีวิตคลัง luteum จะถูกกำหนดโดยระดับของ LH ร่างกายสีเหลืองไม่ใช่เอนทิตีของเซลล์ที่เป็นเนื้อเดียวกัน นอกจากของเซลล์ 2 ประเภท luteal มันมีเซลล์บุผนังหลอดเลือด, ขนาดใหญ่, รบราส์และอื่น ๆ . เซลล์ luteal ขนาดใหญ่ผลิตเปปไทด์ (Relaxin, อุ้ง) และมีการใช้งานมากขึ้นในการ steroidogenesis กิจกรรม aromatase มากขึ้นและการสังเคราะห์ฮอร์โมนที่มีขนาดใหญ่กว่าเซลล์ขนาดเล็ก
จุดสูงสุดของ progesterone จะสังเกตเห็นในวันที่ 8 หลังจากจุดสูงสุดของ LG พบว่าฮอร์โมนโปรเจสเตอโรนและ estradiol ในระยะ luteal จะหลั่งออกมาเป็นระยะ ๆ โดยสัมพันธ์กับพัลส์เอาท์พุตของ LH ด้วยการก่อตัวของสีเหลืองการควบคุมการผลิต inhibin จะผ่าน FSH ไปเป็น LH Ingibin เพิ่มขึ้นเมื่อค่า estradiol เพิ่มขึ้นจนถึงจุดสูงสุดของ LH และยังคงเพิ่มขึ้นหลังจากจุดสูงสุดของ LH แม้ว่าระดับ estrogens จะลดลง แม้ว่ายับยั้งการหลั่งของเอนไซม์ยับยั้งและ estradiol โดยเซลล์ granulosa แต่ก็มีการควบคุมด้วยวิธีต่างๆ การลดลงของ inhibin เมื่อสิ้นสุดระยะ luteal ช่วยเพิ่ม FSH ในรอบถัดไป
ตัวเหลืองอย่างรวดเร็ว - ในวันที่ 9-11 หลังจากการตกไข่ลดลง
กลไกการเสื่อมสภาพไม่ชัดเจนและไม่เกี่ยวข้องกับบทบาทของ estrogen ในไตหรือกลไกการเชื่อมต่อกับ receptor ตามที่เห็นใน endometrium มีคำอธิบายเพิ่มเติมเกี่ยวกับบทบาทของฮอร์โมนเอสโตรเจนที่เกิดจากตัวเหลือง เป็นที่ทราบกันดีว่าสำหรับการสังเคราะห์ตัวรับ progesterone ใน endometrium จำเป็นต้องมี estrogen estrogen อาจมีความจำเป็นสำหรับการเปลี่ยนแปลงที่เกี่ยวข้องกับ progesterone ใน endometrium หลังการตกไข่ การพัฒนาตัวรับ progesterone ที่ไม่เพียงพออันเป็นผลมาจากปริมาณฮอร์โมนเอสโตรเจนที่ไม่เพียงพออาจเป็นกลไกที่ทำให้เกิดภาวะมีบุตรยากและการสูญเสียการตั้งครรภ์ในระยะเริ่มแรกรูปแบบอื่น ๆ ของความด้อยกว่าในระยะ luteal เป็นที่เชื่อกันว่าช่วงชีวิตของร่างกายสีเหลืองถูกตั้งค่าในเวลาของการตกไข่ และมันจะถดถอยอย่างแน่นอนถ้า gonadotropin chorionic ไม่ได้รับการสนับสนุนในการเชื่อมต่อกับการตั้งครรภ์ ดังนั้นการถดถอยของตัวเหลืองทำให้ระดับ estradiol, progesterone และ inhibin ลดลง ลดยับยั้งการยับยั้งการยับยั้งเชื้อ FSH การลดลงของ estradiol และ progesterone ช่วยให้สามารถเรียกคืนการหลั่งของ GnRH ได้อย่างรวดเร็วและลบกลไกการตอบรับจากต่อมใต้สมอง การลด inhibin และ estradiol ร่วมกับการเพิ่ม GnRH ทำให้ความชุกของ FSH มากกว่า LH การเพิ่มขึ้นของ FSH ส่งผลให้มีการเจริญเติบโตของรูขุมขนด้วยการเลือกรูขุมขนที่ตามมาและวัฏจักรใหม่จะเริ่มขึ้นในกรณีที่การตั้งครรภ์ไม่เกิดขึ้น ฮอร์โมนเตียรอยด์มีบทบาทสำคัญในด้านชีววิทยาการสืบพันธุ์และในสรีรวิทยาทั่วไป มีผลต่อระบบหัวใจและหลอดเลือดการเผาผลาญอาหารของกระดูกผิวหนังความเป็นอยู่ทั่วไปของร่างกายและมีบทบาทสำคัญในการตั้งครรภ์ การกระทำของฮอร์โมนสเตียรอยด์สะท้อนกลไกภายในเซลล์และทางพันธุกรรมที่จำเป็นในการถ่ายโอนสัญญาณนอกเซลล์ไปยังนิวเคลียสของเซลล์เพื่อกระตุ้นการตอบสนองทางสรีรวิทยา
เอสโตรเจนกระจายผ่านเยื่อหุ้มเซลล์และผูกกับตัวรับที่อยู่ในนิวเคลียสของเซลล์ เซลล์รับ - สเตียรอยด์จะเชื่อมโยงกับดีเอ็นเอ ในเซลล์เป้าหมายการมีปฏิสัมพันธ์เหล่านี้นำไปสู่การแสดงออกของยีนการสังเคราะห์โปรตีนเพื่อการทำงานเฉพาะของเซลล์และเนื้อเยื่อ