ผู้เชี่ยวชาญทางการแพทย์ของบทความ

สิ่งตีพิมพ์ใหม่
รังไข่
ตรวจสอบล่าสุด: 04.07.2025

เนื้อหา iLive ทั้งหมดได้รับการตรวจสอบทางการแพทย์หรือตรวจสอบข้อเท็จจริงเพื่อให้แน่ใจว่ามีความถูกต้องตามจริงมากที่สุดเท่าที่จะเป็นไปได้
เรามีแนวทางการจัดหาที่เข้มงวดและมีการเชื่อมโยงไปยังเว็บไซต์สื่อที่มีชื่อเสียงสถาบันการวิจัยทางวิชาการและเมื่อใดก็ตามที่เป็นไปได้ โปรดทราบว่าตัวเลขในวงเล็บ ([1], [2], ฯลฯ ) เป็นลิงก์ที่คลิกได้เพื่อการศึกษาเหล่านี้
หากคุณรู้สึกว่าเนื้อหาใด ๆ ของเราไม่ถูกต้องล้าสมัยหรือมีข้อสงสัยอื่น ๆ โปรดเลือกแล้วกด Ctrl + Enter
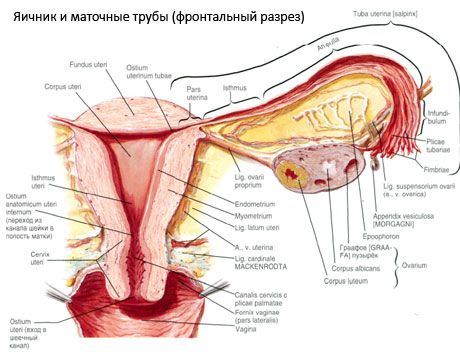
รังไข่ (ovarium; กรีก oophoron) เป็นอวัยวะคู่ เป็นต่อมสืบพันธุ์ของเพศหญิง ตั้งอยู่ในช่องเชิงกรานหลังเอ็นกว้างของมดลูก ในรังไข่ เซลล์สืบพันธุ์ของเพศหญิง (ไข่) จะพัฒนาและโตเต็มที่ และฮอร์โมนเพศหญิงจะถูกสร้างขึ้นเพื่อเข้าสู่กระแสเลือดและน้ำเหลือง รังไข่มีรูปร่างเป็นวงรี แบนเล็กน้อยในทิศทางหน้า-หลัง สีของรังไข่เป็นสีชมพู บนพื้นผิวของรังไข่ของสตรีที่คลอดบุตร จะมองเห็นรอยบุ๋มและรอยแผลเป็น - ร่องรอยของการตกไข่และการเปลี่ยนแปลงของคอร์ปัสลูเตีย รังไข่มีน้ำหนัก 5-8 กรัม ขนาดของรังไข่คือ: ยาว 2.5-5.5 ซม. กว้าง 1.5-3.0 ซม. หนาไม่เกิน 2 ซม. รังไข่มีพื้นผิวอิสระ 2 ด้าน คือ พื้นผิวตรงกลาง (facies medialis) ซึ่งอยู่ติดกับช่องเชิงกรานซึ่งถูกปกคลุมด้วยท่อนำไข่บางส่วน และพื้นผิวด้านข้าง (facies lateralis) ซึ่งอยู่ติดกับผนังด้านข้างของเชิงกรานจนถึงแอ่งที่แสดงออกเล็กน้อย เรียกว่าโพรงรังไข่ โพรงนี้จะอยู่ในมุมระหว่างหลอดเลือดอุ้งเชิงกรานด้านนอกซึ่งปกคลุมด้วยเยื่อบุช่องท้องที่ด้านบนและหลอดเลือดแดงมดลูกและหลอดเลือดอุดตันที่ด้านล่าง ด้านหลังรังไข่ ท่อไตของด้านที่เกี่ยวข้องจะผ่านเข้าไปในช่องท้องจากด้านบนลงด้านล่าง
พื้นผิวของรังไข่จะผ่านเข้าไปในขอบนูน (posterior) (margo liber) ด้านหน้าจะเข้าสู่ขอบของเยื่อบุช่องท้อง (margo mesovaricus) ซึ่งเชื่อมกับแผ่นหลังของเอ็นกว้างของมดลูกด้วยรอยพับสั้นของเยื่อบุช่องท้อง (mesentery of the ovary) ที่ขอบด้านหน้าของอวัยวะนี้จะมีร่องลึกที่เรียกว่า hilum ของรังไข่ (hilum ovarii) ซึ่งหลอดเลือดแดงและเส้นประสาทจะเข้าสู่รังไข่ และหลอดเลือดดำและหลอดน้ำเหลืองจะออกจากรังไข่ รังไข่ยังมีปลายทั้งสองด้าน ได้แก่ ปลายท่อกลมด้านบน (extremitas tubaria) ซึ่งอยู่ตรงข้ามกับท่อนำไข่ และปลายมดลูกด้านล่าง (extremitas utenna) ซึ่งเชื่อมต่อกับมดลูกด้วยเอ็นรังไข่ (lig. ovarii proprium) เอ็นนี้มีลักษณะเป็นเชือกกลมหนาประมาณ 6 มม. ทอดจากปลายมดลูกของรังไข่ไปจนถึงมุมด้านข้างของมดลูก ซึ่งอยู่ระหว่างใบทั้งสองของเอ็นกว้างของมดลูก เอ็นของรังไข่ยังรวมถึงเอ็นแขวนรังไข่ (lig.suspensorium ovarii) ซึ่งเป็นรอยพับของเยื่อบุช่องท้องที่ทอดจากผนังของอุ้งเชิงกรานเล็กไปยังรังไข่ และมีหลอดเลือดของรังไข่และมัดของเส้นใยที่อยู่ภายใน รังไข่จะยึดติดด้วยเมเซนเทอรีสั้น (mesovarium) ซึ่งเป็นการซ้ำซ้อนของเยื่อบุช่องท้องที่ทอดจากใบด้านหลังของเอ็นกว้างของมดลูกไปยังขอบเมเซนเทอรีของรังไข่ รังไข่เองไม่ได้ถูกปกคลุมด้วยเยื่อบุช่องท้อง ขอบรังไข่ที่ใหญ่ที่สุดของท่อนำไข่ติดอยู่ที่ปลายท่อรังไข่ ลักษณะภูมิประเทศของรังไข่ขึ้นอยู่กับตำแหน่งของมดลูก ขนาด (ในระหว่างตั้งครรภ์) รังไข่เป็นอวัยวะที่เคลื่อนไหวได้มากในโพรงเชิงกรานขนาดเล็ก
 [
[ หลอดเลือดและเส้นประสาทของรังไข่
เลือดที่ไปเลี้ยงรังไข่มาจาก aa. et vv. ovaricae et uterinae หลอดเลือดแดงรังไข่ทั้งสองเส้น (aa. ovaricae dextra et sinistra) มีจุดเริ่มต้นจากพื้นผิวด้านหน้าของหลอดเลือดแดงใหญ่ใต้หลอดเลือดแดงไต หลอดเลือดแดงด้านขวามักจะมาจากหลอดเลือดแดงไต และหลอดเลือดแดงด้านซ้ายจะมาจากหลอดเลือดแดงไต หลอดเลือดแดงรังไข่แต่ละเส้นจะมุ่งลงและไปด้านข้างตามพื้นผิวด้านหน้าของกล้ามเนื้อ psoas major โดยจะข้ามท่อไตด้านหน้า (โดยแตกแขนงออกไป) หลอดเลือดอุ้งเชิงกรานภายนอก เส้นขอบเขต และเข้าสู่ช่องเชิงกราน โดยตั้งอยู่ในเอ็นยึดรังไข่ หลอดเลือดแดงรังไข่จะวิ่งตามทิศทางตรงกลางระหว่างใบของเอ็นยึดมดลูกใต้ท่อนำไข่ โดยแตกแขนงออกไป จากนั้นจึงเข้าสู่เยื่อหุ้มของรังไข่ มันเข้าสู่ช่องของรังไข่
กิ่งก้านของหลอดเลือดแดงรังไข่เชื่อมต่อกับกิ่งก้านของหลอดเลือดแดงมดลูกอย่างกว้างขวาง การไหลของเลือดดำจากรังไข่จะไหลเข้าสู่กลุ่มหลอดเลือดดำของรังไข่เป็นหลัก ซึ่งตั้งอยู่ในบริเวณของไฮลัมของรังไข่ จากจุดนี้ เลือดจะไหลออกในสองทิศทาง คือ ผ่านหลอดเลือดดำของมดลูกและหลอดเลือดดำของรังไข่ หลอดเลือดดำรังไข่ด้านขวามีลิ้นและไหลเข้าสู่ vena cava inferior หลอดเลือดดำรังไข่ด้านซ้ายไหลเข้าสู่หลอดเลือดดำของไตด้านซ้าย แม้ว่าจะไม่มีลิ้นก็ตาม
การระบายน้ำเหลืองจากรังไข่เกิดขึ้นผ่านหลอดน้ำเหลือง โดยเฉพาะอย่างยิ่งในบริเวณประตูอวัยวะซึ่งแยกกลุ่มหลอดน้ำเหลืองใต้รังไข่ จากนั้นน้ำเหลืองจะถูกระบายไปตามหลอดน้ำเหลืองของรังไข่ไปยังต่อมน้ำเหลืองข้างหัวใจ
การส่งสัญญาณประสาทไปยังรังไข่
ระบบซิมพาเทติก – มาจากใยประสาทหลังปมประสาทจากช่องท้องส่วนบน (solar plexuses) ช่องท้องส่วนบน (superior mesenteric) และช่องท้องส่วนล่าง (hypogastric plexuses) ระบบพาราซิมพาเทติก – มาจากเส้นประสาทที่กระดูกสันหลังส่วนใน
โครงสร้างของรังไข่
พื้นผิวของรังไข่ถูกปกคลุมด้วยเยื่อบุผิวเจริญชั้นเดียว ด้านล่างมีชั้นโปรตีนเนื้อเยื่อเกี่ยวพันหนาแน่น (tunica albuginea) เนื้อเยื่อเกี่ยวพันของรังไข่สร้างเนื้อเยื่อเกี่ยวพัน (stroma ovarii) ซึ่งอุดมไปด้วยเส้นใยยืดหยุ่น สารในรังไข่หรือพาเรนไคมาแบ่งออกเป็นชั้นนอกและชั้นใน ชั้นในซึ่งอยู่ตรงกลางรังไข่ ใกล้กับประตูรังไข่ เรียกว่าเมดัลลา (medulla ovarii) ในชั้นนี้ มีหลอดเลือดและน้ำเหลืองและเส้นประสาทจำนวนมากในเนื้อเยื่อเกี่ยวพันหลวม ชั้นนอกของรังไข่ซึ่งเรียกว่าคอร์เทกซ์ (cortex ovarii) มีความหนาแน่นมากกว่า ประกอบด้วยเนื้อเยื่อเกี่ยวพันจำนวนมาก ซึ่งอยู่ในรูขุมขนปฐมภูมิของรังไข่ที่เจริญเต็มที่ (folliculi ovarici primarii), รูขุมขนรอง (ตุ่ม) (folliculi ovarici secundarii, s.vesiculosi) เช่นเดียวกับรูขุมขนที่โตเต็มวัย รูขุมขน Graafian (folliculi ovarici maturis) เช่นเดียวกับร่างกายสีเหลืองและ atretic
แต่ละฟอลลิเคิลจะมีเซลล์สืบพันธุ์เพศเมียหรือโอโอไซต์ (ovocytus) เซลล์ไข่มีเส้นผ่านศูนย์กลางประมาณ 150 ไมโครเมตร มีลักษณะกลม มีนิวเคลียส ไซโทพลาซึมจำนวนมาก ซึ่งนอกจากออร์แกเนลล์ของเซลล์แล้ว ยังมีโปรตีนและลิพิด (ไข่แดง) ไกลโคเจน ซึ่งจำเป็นต่อการสร้างอาหารให้กับเซลล์ไข่ โดยปกติแล้วไข่จะใช้สารอาหารที่สะสมภายใน 12-24 ชั่วโมงหลังตกไข่ หากไม่เกิดการปฏิสนธิ ไข่ก็จะตาย
ไข่ของมนุษย์มีเยื่อหุ้ม 2 ชั้น ด้านในคือไซโตเลมมา ซึ่งเป็นเยื่อหุ้มไซโตพลาสซึมของไข่ ด้านนอกของไซโตเลมมาคือชั้นของเซลล์ที่เรียกว่า ฟอลลิคูลาร์ ซึ่งทำหน้าที่ปกป้องไข่และทำหน้าที่สร้างฮอร์โมน โดยเซลล์เหล่านี้จะหลั่งฮอร์โมนเอสโตรเจน
ตำแหน่งทางสรีรวิทยาของมดลูก ท่อนำไข่ และรังไข่เกิดจากกลไกการแขวน การตรึง และการรองรับที่เชื่อมเยื่อบุช่องท้อง เอ็น และเนื้อเยื่อเชิงกรานเข้าด้วยกัน กลไกการแขวนจะแสดงเป็นโครงสร้างคู่ ซึ่งประกอบด้วยเอ็นกลมและเอ็นกว้างของมดลูก เอ็นที่เหมาะสม และเอ็นแขวนของรังไข่ เอ็นกว้างของมดลูก เอ็นที่เหมาะสมและเอ็นแขวนของรังไข่ ยึดมดลูกไว้ในตำแหน่งตรงกลาง เอ็นกลมดึงก้นมดลูกไปข้างหน้าและเอียงตามสรีรวิทยา
อุปกรณ์ยึด (anchoring) ช่วยให้มดลูกอยู่ในตำแหน่งตรงกลางของอุ้งเชิงกรานเล็ก และทำให้มดลูกไม่สามารถเคลื่อนไปด้านข้างหรือไปข้างหน้าหรือข้างหลังได้ แต่เนื่องจากอุปกรณ์ยึดมดลูกออกจากมดลูกในส่วนล่าง จึงทำให้มดลูกเอียงไปในทิศทางต่างๆ ได้ อุปกรณ์ยึดมดลูกประกอบด้วยเอ็นที่อยู่ในเนื้อเยื่อที่หลวมของอุ้งเชิงกรานและทอดยาวจากส่วนล่างของมดลูกไปยังผนังด้านข้าง ด้านหน้า และด้านหลังของอุ้งเชิงกราน ได้แก่ เอ็น sacro-magic, cardinal, uterovesical และ vesicopubic
นอกจากเมโซวาเรียมแล้ว ยังมีเอ็นรังไข่อีกดังต่อไปนี้:
- เอ็นยึดรังไข่ ซึ่งก่อนหน้านี้เรียกว่าเอ็นยึดกระดูกเชิงกราน (infundibulopelvic ligament) เป็นรอยพับของเยื่อบุช่องท้องที่มีหลอดเลือด (a. et v. ovarica) และหลอดน้ำเหลืองและเส้นประสาทของรังไข่ผ่านเข้าไป ยืดระหว่างผนังด้านข้างของอุ้งเชิงกราน พังผืดเอว (ในบริเวณที่แบ่งหลอดเลือดแดงอุ้งเชิงกรานส่วนนอกและส่วนใน) และปลายด้านบน (ท่อนำไข่) ของรังไข่
- เอ็นที่เหมาะสมของรังไข่จะผ่านระหว่างใบของเอ็นกว้างของมดลูก ใกล้กับใบหลัง และเชื่อมปลายด้านล่างของรังไข่กับขอบด้านข้างของมดลูก เอ็นที่เหมาะสมของรังไข่จะติดกับมดลูกระหว่างจุดเริ่มต้นของท่อนำไข่และเอ็นกลม ด้านหลังและด้านบนของเอ็นกลม ในความหนาของเอ็นจะผ่าน rr. ovarii ซึ่งเป็นกิ่งปลายสุดของหลอดเลือดแดงมดลูก
- เอ็นไส้ติ่ง-รังไข่ เอ็นนี้ทอดยาวจากเยื่อหุ้มไส้ติ่งของไส้ติ่งไปจนถึงรังไข่ด้านขวาหรือเอ็นกว้างของมดลูกในรูปของรอยพับของเยื่อบุช่องท้อง เอ็นนี้มีลักษณะไม่คงตัวและพบในผู้หญิงประมาณ 1/2 - 1/3
อุปกรณ์สนับสนุนประกอบด้วยกล้ามเนื้อและพังผืดของพื้นเชิงกราน ซึ่งแบ่งออกเป็นชั้นล่าง ชั้นกลาง และชั้นบน (ชั้นใน)
ชั้นกล้ามเนื้อด้านบน (ชั้นใน) ถือเป็นชั้นกล้ามเนื้อคู่ที่ทำหน้าที่ยกทวารหนัก ชั้นกล้ามเนื้อนี้ประกอบด้วยมัดกล้ามเนื้อที่แผ่ขยายจากกระดูกก้นกบไปยังกระดูกเชิงกรานใน 3 ทิศทาง (พูโบค็อกไซเจียส อิลิโอค็อกไซเจียส และอิสคิโอค็อกไซเจียส) ชั้นกล้ามเนื้อนี้เรียกอีกอย่างว่ากะบังลมเชิงกราน
ชั้นกลางของกล้ามเนื้อตั้งอยู่ระหว่างซิมฟิซิส กระดูกหัวหน่าว และกระดูกก้นกบ ชั้นกลางของกล้ามเนื้อ - กะบังลมทางเดินปัสสาวะและอวัยวะสืบพันธุ์ - ครอบครองครึ่งด้านหน้าของช่องทางอุ้งเชิงกราน ซึ่งเป็นทางผ่านของท่อปัสสาวะและช่องคลอด ในส่วนหน้าระหว่างแผ่นของกล้ามเนื้อมีมัดกล้ามเนื้อที่สร้างหูรูดภายนอกของท่อปัสสาวะ ในส่วนหลังมีมัดกล้ามเนื้อที่ไปในทิศทางขวาง - กล้ามเนื้อขวางลึกของฝีเย็บ
ชั้นล่าง (ชั้นนอก) ของกล้ามเนื้ออุ้งเชิงกรานประกอบด้วยกล้ามเนื้อผิวเผินซึ่งมีรูปร่างคล้ายเลข 8 ได้แก่ กล้ามเนื้อบัลโบคาเวอร์นัส กล้ามเนื้ออิสคิโอคาเวอร์นัส กล้ามเนื้อหูรูดทวารหนักภายนอก และกล้ามเนื้อขวางของฝีเย็บผิวเผิน
การเกิดรังไข่
กระบวนการเจริญเติบโตของฟอลลิเคิลและภาวะแข็งเกร็งเริ่มต้นเมื่ออายุครรภ์ได้ 20 สัปดาห์ และเมื่อถึงเวลาคลอด จะมีไข่เหลืออยู่ในรังไข่ของเด็กผู้หญิงมากถึง 2 ล้านฟอง เมื่อถึงประจำเดือน จำนวนไข่จะลดลงเหลือ 300,000 ฟอง ตลอดช่วงชีวิตการสืบพันธุ์ ฟอลลิเคิลไม่เกิน 500 ฟอลลิเคิลจะเจริญเติบโตเต็มที่และตกไข่ การเจริญเติบโตในช่วงแรกของฟอลลิเคิลไม่ได้ขึ้นอยู่กับการกระตุ้นของ FSH แต่จะมีจำกัด และภาวะแข็งเกร็งจะเกิดขึ้นอย่างรวดเร็ว เชื่อกันว่าแทนที่จะใช้ฮอร์โมนสเตียรอยด์ เปปไทด์ออโตไครน์/พาราไครน์ในท้องถิ่นจะเป็นตัวควบคุมหลักในการเจริญเติบโตและภาวะแข็งเกร็งของฟอลลิเคิลหลัก เชื่อกันว่ากระบวนการเจริญเติบโตของฟอลลิเคิลและภาวะแข็งเกร็งจะไม่ถูกขัดจังหวะด้วยกระบวนการทางสรีรวิทยาใดๆ กระบวนการนี้ดำเนินต่อไปในทุกช่วงวัย รวมทั้งช่วงที่อยู่ในครรภ์และวัยหมดประจำเดือน และถูกขัดจังหวะด้วยการตั้งครรภ์ การตกไข่ และการไม่ตกไข่ กลไกที่กระตุ้นการเจริญเติบโตของฟอลลิเคิลและจำนวนฟอลลิเคิลในแต่ละรอบยังคงไม่ชัดเจน
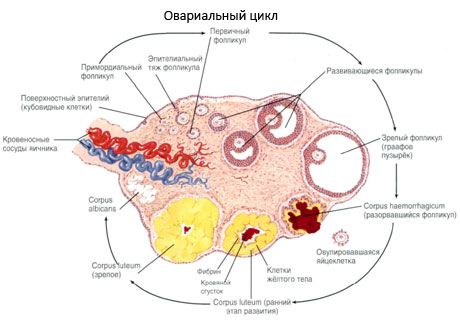
ในระหว่างการพัฒนา ฟอลลิเคิลจะผ่านขั้นตอนการพัฒนาหลายขั้นตอน เซลล์สืบพันธุ์ดั้งเดิมมีต้นกำเนิดจากเอนโดเดิร์มของถุงไข่แดง อัลลันทอยส์ และอพยพไปยังบริเวณอวัยวะเพศของตัวอ่อนเมื่ออายุครรภ์ได้ 5-6 สัปดาห์ เนื่องมาจากการแบ่งตัวแบบไมโทซิสอย่างรวดเร็ว ซึ่งดำเนินต่อไปตั้งแต่ 6-8 สัปดาห์ถึง 16-20 สัปดาห์ของการตั้งครรภ์ เซลล์ไข่มากถึง 6-7 ล้านเซลล์จะก่อตัวในรังไข่ของตัวอ่อน ซึ่งล้อมรอบด้วยเซลล์เกรนูลูซาชั้นบางๆ
รูเปิดของไข่ - ไข่ถูกล้อมรอบด้วยเยื่อหุ้ม (zona pellucida) เซลล์แกรนูโลซาที่ล้อมรอบไข่จะเริ่มขยายตัว การเจริญเติบโตขึ้นอยู่กับฮอร์โมนโกนาโดโทรปินและสัมพันธ์กับระดับของเอสโตรเจน เซลล์แกรนูโลซาเป็นเป้าหมายของ FSH ในระยะรูเปิดของไข่ เซลล์แกรนูโลซาสามารถสังเคราะห์สเตียรอยด์ได้สามกลุ่ม โดยส่วนใหญ่กระตุ้นการทำงานของอะโรมาเตส ซึ่งเป็นเอนไซม์หลักที่เปลี่ยนแอนโดรเจนเป็นเอสตราไดออล เชื่อกันว่าเอสตราไดออลสามารถเพิ่มจำนวนตัวรับของตัวเองได้ โดยออกฤทธิ์โดยตรงต่อเซลล์แกรนูโลซาโดยไม่ขึ้นกับ FSH เอสตราไดออลถือเป็นปัจจัยพาราไครน์ที่ช่วยเพิ่มผลของ FSH รวมถึงกระตุ้นกระบวนการอะโรมาเตชันด้วย
ตัวรับ FSH ปรากฏบนเยื่อหุ้มเซลล์เกรนูลูซาทันทีที่รูขุมขนเริ่มเติบโต การลดลงหรือเพิ่มขึ้นของ FSH ส่งผลให้จำนวนตัวรับเปลี่ยนแปลงไป การทำงานของ FSH นี้ได้รับการปรับเปลี่ยนโดยปัจจัยการเจริญเติบโต FSH ออกฤทธิ์ผ่านโปรตีน G หรือระบบอะดีไนเลตไซเคลส แม้ว่าการสร้างสเตียรอยด์ในรูขุมขนจะถูกควบคุมโดย FSH เป็นหลัก แต่ก็มีปัจจัยหลายประการที่เกี่ยวข้องในกระบวนการนี้ ได้แก่ ช่องไอออน ตัวรับไทโรซีนไคเนส ระบบฟอสโฟไลเปสของตัวส่งสารที่สอง
บทบาทของแอนโดรเจนในการพัฒนาของรูขุมขนในระยะเริ่มต้นนั้นมีความซับซ้อน เซลล์กรานูโลซามีตัวรับแอนโดรเจน ซึ่งไม่เพียงแต่เป็นสารตั้งต้นสำหรับอะโรมาไทเซชันที่เหนี่ยวนำโดย FSH เป็นเอสโตรเจนเท่านั้น แต่ยังสามารถเพิ่มอะโรมาไทเซชันในความเข้มข้นต่ำได้อีกด้วย เมื่อระดับแอนโดรเจนเพิ่มขึ้น เซลล์กรานูโลซาก่อนแอนทรัลจะเลือกเส้นทางอะโรมาไทเซชันไปยังเอสโตรเจนแทนเส้นทางอะโรมาไทเซชัน แต่เลือกเส้นทางแอนโดรเจนที่ง่ายกว่าผ่าน 5a-reductase เพื่อเปลี่ยนเป็นแอนโดรเจนที่ไม่สามารถเปลี่ยนเป็นเอสโตรเจนได้ จึงยับยั้งกิจกรรมของอะโรมาไทเซชัน กระบวนการนี้ยังยับยั้งการสร้างตัวรับ FSH และ LH จึงหยุดการพัฒนาของรูขุมขน
กระบวนการอะโรมาไทเซชัน ฟอลลิเคิลที่มีระดับแอนโดรเจนสูงจะเข้าสู่กระบวนการอะเทรเซีย การเจริญเติบโตและการพัฒนาของฟอลลิเคิลขึ้นอยู่กับความสามารถในการเปลี่ยนแอนโดรเจนเป็นเอสโตรเจน
ในกรณีที่มี FSH สารหลักของของเหลวในรูขุมขนจะเป็นเอสโตรเจน ในกรณีที่ไม่มี FSH จะเป็นแอนโดรเจน LH มักจะไม่มีอยู่ในของเหลวในรูขุมขนจนถึงกลางรอบเดือน ทันทีที่ระดับ LH เพิ่มขึ้น กิจกรรมไมโทซิสของเซลล์เกรนูลูซาจะลดลง การเปลี่ยนแปลงทางเสื่อมเกิดขึ้น และระดับแอนโดรเจนในรูขุมขนจะเพิ่มขึ้น ระดับของสเตียรอยด์ในของเหลวในรูขุมขนสูงกว่าในพลาสมา และสะท้อนถึงกิจกรรมการทำงานของเซลล์รังไข่: เซลล์เกรนูลูซาและเซลล์ธีคา หากเป้าหมายเดียวของ FSH คือเซลล์เกรนูลูซา LH ก็มีเป้าหมายมากมาย ได้แก่ เซลล์ธีคา เซลล์สโตรมาและลูทีน และเซลล์เกรนูลูซา ทั้งเซลล์เกรนูลูซาและเซลล์ธีคาต่างก็มีความสามารถในการสร้างสเตียรอยด์ แต่กิจกรรมของอะโรมาเทสจะโดดเด่นกว่าในเซลล์เกรนูลูซา
ในการตอบสนองต่อ LH เซลล์ theca จะผลิตแอนโดรเจน ซึ่งจะถูกเซลล์เกรนูลูซาแปลงให้เป็นเอสโตรเจนผ่านการสร้างอะโรมาไทเซชันที่เหนี่ยวนำโดย FSH
ขณะที่รูขุมขนเจริญเติบโต เซลล์ theca จะเริ่มแสดงยีนสำหรับตัวรับ LH, P450 sec และ 3beta-hydroxysteroid dehydrogenase, อินซูลินไลค์โกรทแฟกเตอร์ (IGF-1) ร่วมกับ LH จะเพิ่มการแสดงออกของยีน แต่จะไม่กระตุ้นการสร้างสเตียรอยด์
การสร้างสเตียรอยด์ในรังไข่ขึ้นอยู่กับ LH เสมอ เมื่อฟอลลิเคิลเติบโต เซลล์ theca จะแสดงเอนไซม์ P450c17 ซึ่งสร้างแอนโดรเจนจากคอเลสเตอรอล เซลล์ Granulosa ไม่มีเอนไซม์นี้และต้องอาศัยเซลล์ theca เพื่อสร้างเอสโตรเจนจากแอนโดรเจน ซึ่งแตกต่างจากการสร้างสเตียรอยด์ การสร้างฟอลลิเคิลขึ้นอยู่กับ FSH เมื่อฟอลลิเคิลเติบโตและระดับเอสโตรเจนเพิ่มขึ้น กลไกการป้อนกลับจะถูกเปิดใช้งาน โดยการผลิต FSH จะถูกยับยั้ง ซึ่งส่งผลให้กิจกรรมอะโรมาเตสของฟอลลิเคิลลดลง และสุดท้ายจะเกิดภาวะฟอลลิเคิลแอเทรเซียผ่านอะพอพโทซิส (การตายของเซลล์ตามโปรแกรม)
กลไกการป้อนกลับของเอสโตรเจนและ FSH ยับยั้งการพัฒนาของฟอลลิเคิลที่เริ่มเติบโตแล้ว แต่ไม่ใช่ฟอลลิเคิลหลัก ฟอลลิเคิลหลักมีตัวรับ FSH มากกว่า ซึ่งสนับสนุนการแพร่กระจายของเซลล์เกรนูลูซาและการเปลี่ยนแอนโดรเจนเป็นเอสโตรเจน นอกจากนี้ เส้นทางพาราไครน์และออโตไครน์ยังทำหน้าที่เป็นตัวประสานงานที่สำคัญในการพัฒนาฟอลลิเคิลแอนทรัลอีกด้วย
ตัวควบคุมออโตไครน์/พาราไครน์ประกอบด้วยเปปไทด์ (อินฮิบิน แอคติวิน ฟอลลิสแตติน) ซึ่งสังเคราะห์โดยเซลล์เกรนูลูซาเพื่อตอบสนองต่อ FSH และเข้าสู่ของเหลวในรูขุมขน อินฮิบินช่วยลดการหลั่ง FSH แอคติวินกระตุ้นการปล่อย FSH จากต่อมใต้สมองและเพิ่มการทำงานของ FSH ในรังไข่ ฟอลลิสแตตินยับยั้งการทำงานของ FSH โดยอาจจับกับแอคติวิน หลังจากการตกไข่และการพัฒนาของคอร์ปัสลูเทียม อินฮิบินจะอยู่ภายใต้การควบคุมของ LH
การเจริญเติบโตและการแบ่งตัวของเซลล์รังไข่ได้รับอิทธิพลจากอินซูลินไลค์โกรทแฟกเตอร์ (IGE) IGF-1 ออกฤทธิ์กับเซลล์เกรนูลูซา ทำให้มีไซคลิกอะดีโนซีนโมโนฟอสเฟต (cAMP) โปรเจสเตอโรน ออกซิโทซิน โปรตีโอไกลแคน และอินฮิบินเพิ่มขึ้น
IGF-1 ออกฤทธิ์ต่อเซลล์ธีคา ทำให้มีการผลิตแอนโดรเจนเพิ่มขึ้น เซลล์ธีคาจะผลิตเนื้องอกเนโครซิสแฟกเตอร์ (TNF) และเอพิเดอร์มัลโกรทแฟกเตอร์ (EGF) ซึ่งควบคุมโดย FSH เช่นกัน
EGF ช่วยกระตุ้นการแบ่งตัวของเซลล์เม็ดเลือดขาว IGF-2 เป็นปัจจัยหลักในการเจริญเติบโตของของเหลวในรูขุมขน นอกจากนี้ยังพบ IGF-1, TNF-α, TNF-3 และ EGF อีกด้วย
การหยุดชะงักของการควบคุมพาราไครน์และ/หรือออโตไครน์ของการทำงานของรังไข่อาจมีบทบาทในความผิดปกติของการตกไข่และการพัฒนาของรังไข่ที่มีถุงน้ำจำนวนมาก
เมื่อรูขุมขนขยายใหญ่ขึ้น ปริมาณเอสโตรเจนในของเหลวของรูขุมขนก็จะเพิ่มขึ้น เมื่อรูขุมขนขยายถึงจุดสูงสุด ตัวรับ LH จะปรากฏขึ้นบนเซลล์เกรนูลูซา เซลล์เกรนูลูซาจะมีลูทีนเพิ่มขึ้น และการผลิตโปรเจสเตอโรนจะเพิ่มขึ้น ดังนั้น ในช่วงก่อนการตกไข่ การผลิตเอสโตรเจนที่เพิ่มขึ้นจะทำให้เกิดตัวรับ LH ขึ้น ซึ่ง LH จะทำให้เซลล์เกรนูลูซามีลูทีนเพิ่มขึ้นและเกิดการผลิตโปรเจสเตอโรน การเพิ่มขึ้นของโปรเจสเตอโรนจะทำให้ระดับเอสโตรเจนลดลง ซึ่งเห็นได้ชัดว่าทำให้เกิด FSH พุ่งสูงเป็นครั้งที่สองในช่วงกลางรอบเดือน
เชื่อกันว่าการตกไข่จะเกิดขึ้น 10-12 ชั่วโมงหลังจากระดับสูงสุดของฮอร์โมน LH และ 24-36 ชั่วโมงหลังจากระดับสูงสุดของฮอร์โมนเอสตราไดออล เชื่อกันว่าฮอร์โมน LH กระตุ้นการแบ่งตัวของไข่ การเกิดลูทีนในเซลล์เกรนูลูซา และการสังเคราะห์โปรเจสเตอโรนและพรอสตาแกลนดินในรูขุมขน
โปรเจสเตอโรนช่วยเพิ่มการทำงานของเอนไซม์โปรตีโอไลติก ซึ่งร่วมกับพรอสตาแกลนดินมีส่วนในการแตกของผนังรูขุมขน โปรเจสเตอโรนกระตุ้นจุดสูงสุดของ FSH ช่วยให้โอโอไซต์ออกจากรูขุมขนได้โดยเปลี่ยนพลาสมินเจนเป็นเอนไซม์โปรตีโอไลติกพลาสมิน และให้ตัวรับ LH ในจำนวนที่เพียงพอสำหรับการพัฒนาปกติของระยะลูเตียล
ภายใน 3 วันหลังจากการตกไข่ เซลล์กรานูโลซาจะมีขนาดใหญ่ขึ้น และช่องว่างที่มีลักษณะเฉพาะซึ่งเต็มไปด้วยเม็ดสี ลูทีน จะปรากฏขึ้นในเซลล์เหล่านี้ เซลล์ธีคาลูเตียลจะแยกความแตกต่างจากธีคาและสโตรมา และกลายเป็นส่วนหนึ่งของคอร์ปัสลูเตียม เส้นเลือดฝอยที่เจาะเข้าไปในคอร์ปัสลูเตียมจะพัฒนาอย่างรวดเร็วภายใต้อิทธิพลของปัจจัยการสร้างหลอดเลือดใหม่ และด้วยการสร้างหลอดเลือดที่ดีขึ้น การผลิตโปรเจสเตอโรนและเอสโตรเจนจะเพิ่มขึ้น กิจกรรมของการสร้างสเตียรอยด์และอายุขัยของคอร์ปัสลูเตียมจะถูกกำหนดโดยระดับของ LH คอร์ปัสลูเตียมไม่ใช่การก่อตัวของเซลล์ที่เป็นเนื้อเดียวกัน นอกจากเซลล์ลูเตียล 2 ประเภทแล้ว ยังมีเซลล์เยื่อบุผนังหลอดเลือด แมคโครฟาจ ไฟโบรบลาสต์ ฯลฯ อีกด้วย เซลล์ลูเตียลขนาดใหญ่จะผลิตเปปไทด์ (รีแลกซิน ออกซิโทซิน) และมีฤทธิ์ในการสร้างสเตียรอยด์มากกว่า โดยมีกิจกรรมอะโรมาเทสมากกว่าและมีการสังเคราะห์โปรเจสเตอโรนมากกว่าเซลล์ขนาดเล็ก
ระดับโปรเจสเตอโรนสูงสุดจะเกิดขึ้นในวันที่ 8 หลังจากระดับ LH สูงสุด โดยจะสังเกตได้ว่าระดับโปรเจสเตอโรนและเอสตราไดออลจะถูกหลั่งออกมาเป็นระยะๆ ในระยะลูเทียล ซึ่งสัมพันธ์กับระดับ LH ที่เต้นเป็นจังหวะ เมื่อมีการสร้างคอร์ปัสลูเทียม การควบคุมการผลิตอินฮิบินจะผ่านจาก FSH ไปยัง LH อินฮิบินจะเพิ่มขึ้นตามระดับเอสตราไดออลที่เพิ่มขึ้นก่อนถึงระดับ LH สูงสุด และจะเพิ่มขึ้นต่อไปหลังจากระดับ LH สูงสุด แม้ว่าระดับเอสโตรเจนจะลดลงก็ตาม แม้ว่าเซลล์กรานูโลซาจะหลั่งอินฮิบินและเอสตราไดออล แต่ทั้งสองอย่างนี้จะถูกควบคุมโดยทางเดินที่แตกต่างกัน การลดลงของอินฮิบินในตอนท้ายของระยะลูเทียลจะส่งผลต่อระดับ FSH ที่เพิ่มขึ้นในรอบต่อไป
คอร์ปัสลูเทียมจะลดลงอย่างรวดเร็วมาก - ในวันที่ 9-11 หลังจากการตกไข่
กลไกการเสื่อมสภาพยังไม่ชัดเจนและไม่เกี่ยวข้องกับบทบาทการสลายลูเตียมของเอสโตรเจนหรือกลไกที่เกี่ยวข้องกับตัวรับตามที่สังเกตในเยื่อบุโพรงมดลูก มีคำอธิบายอื่นสำหรับบทบาทของเอสโตรเจนที่ผลิตโดยคอร์ปัสลูเตียม เป็นที่ทราบกันดีว่าเอสโตรเจนจำเป็นสำหรับการสังเคราะห์ตัวรับโปรเจสเตอโรนในเยื่อบุโพรงมดลูก เอสโตรเจนในระยะลูเตียมอาจจำเป็นสำหรับการเปลี่ยนแปลงที่เกี่ยวข้องกับโปรเจสเตอโรนในเยื่อบุโพรงมดลูกหลังจากการตกไข่ การพัฒนาตัวรับโปรเจสเตอโรนที่ไม่เพียงพออันเป็นผลมาจากระดับเอสโตรเจนที่ไม่เพียงพอ อาจเป็นกลไกเพิ่มเติมของภาวะมีบุตรยากและการแท้งบุตรในระยะแรก ซึ่งเป็นรูปแบบอื่นของภาวะขาดเฟสลูเตียม เชื่อกันว่าอายุขัยของคอร์ปัสลูเตียมจะเกิดขึ้นในช่วงเวลาที่มีการตกไข่ และแน่นอนว่าจะถดถอยลงหากไม่ได้รับการสนับสนุนจากฮอร์โมนโกนาโดโทรปินในมนุษย์เนื่องจากการตั้งครรภ์ ดังนั้น การถดถอยของคอร์ปัสลูเทียมจะนำไปสู่การลดลงของระดับเอสตราไดออล โปรเจสเตอโรน และอินฮิบิน การลดลงของอินฮิบินจะขจัดผลกดการทำงานของ FSH การลดลงของเอสตราไดออลและโปรเจสเตอโรนทำให้การหลั่ง GnRH กลับมาเป็นปกติได้อย่างรวดเร็วและกลไกการป้อนกลับจากต่อมใต้สมองจะถูกกำจัด การลดลงของอินฮิบินและเอสตราไดออล รวมถึงการเพิ่มขึ้นของ GnRH จะทำให้เกิด FSH มากกว่า LH การเพิ่มขึ้นของ FSH นำไปสู่การเจริญเติบโตของฟอลลิเคิล ตามด้วยการคัดเลือกฟอลลิเคิลที่โดดเด่น และวงจรใหม่จะเริ่มขึ้นหากไม่เกิดการตั้งครรภ์ ฮอร์โมนสเตียรอยด์มีบทบาทสำคัญในชีววิทยาการสืบพันธุ์และสรีรวิทยาทั่วไป ฮอร์โมนสเตียรอยด์กำหนดลักษณะทางฟีโนไทป์ของมนุษย์ ส่งผลต่อระบบหัวใจและหลอดเลือด การเผาผลาญของกระดูก ผิวหนัง ความเป็นอยู่โดยรวม และมีบทบาทสำคัญในการตั้งครรภ์ การทำงานของฮอร์โมนสเตียรอยด์สะท้อนกลไกภายในเซลล์และทางพันธุกรรมที่จำเป็นในการส่งสัญญาณภายนอกเซลล์ไปยังนิวเคลียสของเซลล์เพื่อก่อให้เกิดการตอบสนองทางสรีรวิทยา
เอสโตรเจนแทรกซึมผ่านเยื่อหุ้มเซลล์และจับกับตัวรับที่อยู่ในนิวเคลียสของเซลล์ จากนั้นคอมเพล็กซ์ตัวรับ-สเตียรอยด์จะจับกับดีเอ็นเอ ในเซลล์เป้าหมาย ปฏิกิริยาเหล่านี้จะนำไปสู่การแสดงออกของยีน การสังเคราะห์โปรตีน และการทำงานของเซลล์และเนื้อเยื่อเฉพาะ